Lekcija: Sporta treniņu bioķīmiskie pamati. Bioķīmisko procesu dinamika organismā muskuļu darba laikā Muskuļu darba bioķīmija
Kā sportista ķermenis pielāgojas intensīvai muskuļu darbībai?
Dziļās funkcionālās izmaiņas organismā, kas radušās, pielāgojot to paaugstinātai muskuļu aktivitātei, pēta sporta fizioloģija. Taču to pamatā ir bioķīmiskās izmaiņas audu un orgānu vielmaiņā un galu galā arī organismā kopumā. Tomēr mēs vispārīgākajā veidā apsvērsim galvenās izmaiņas, kas notiek treniņu ietekmē tikai muskuļos.
Muskuļu bioķīmiskā pārstrukturēšana treniņu ietekmē ir balstīta uz izdevumu un muskuļu funkcionālo un enerģijas rezervju atjaunošanas procesu savstarpējo atkarību. Kā jau sapratāt no iepriekšējā, muskuļu aktivitātes laikā notiek intensīva ATP šķelšanās un attiecīgi intensīvi tiek patērētas citas vielas. Muskuļos tas ir kreatīna fosfāts, glikogēns, lipīdi, aknās glikogēns sadalās, veidojot cukuru, kas kopā ar asinīm tiek pārnests uz strādājošiem muskuļiem, sirdi un smadzenēm; tauki tiek sadalīti un taukskābes oksidējas. Tajā pašā laikā organismā uzkrājas vielmaiņas produkti - fosforskābe un pienskābe, ketonvielas, oglekļa dioksīds. Daļēji tos organisms pazaudē un daļēji atkal izmanto, iesaistoties vielmaiņā. Muskuļu darbību pavada daudzu enzīmu aktivitātes palielināšanās, un tāpēc sākas izlietoto vielu sintēze. ATP, kreatīna fosfāta un glikogēna resintēze jau ir iespējama darba laikā, taču līdz ar to notiek intensīva šo vielu sadalīšanās. Tāpēc to saturs muskuļos darba laikā nekad nesasniedz oriģinālu.
Atpūtas periodā, kad intensīva enerģijas avotu sadalīšana apstājas, resintēzes procesi iegūst skaidru pārsvaru un notiek ne tikai iztērētā atjaunošana (kompensācija), bet arī superatgūšana (superkompensācija), kas pārsniedz sākotnējo. līmenī. Šo modeli sauc par "superkompensācijas likumu".
Superkompensācijas fenomena būtība.
Sporta bioķīmijā ir pētītas šī procesa likumsakarības. Piemēram, ir konstatēts, ka, ja muskuļos, aknās un citos orgānos notiek intensīva vielas iztērēšana, jo ātrāk notiek resintēze un jo izteiktāka ir pārmērīgas atveseļošanās parādība. Piemēram, pēc īslaicīga intensīva darba glikogēna līmeņa paaugstināšanās muskuļos, kas pārsniedz sākotnējo, notiek pēc 1 stundas atpūtas, un pēc 12 stundām tas atgriežas sākotnējā, galīgajā līmenī. Ilgstoši strādājot, superkompensācija notiek tikai pēc 12 stundām, bet paaugstināts glikogēna līmenis muskuļos saglabājas ilgāk par trim dienām. Tas ir iespējams tikai pateicoties augstajai enzīmu aktivitātei un to pastiprinātai sintēzei.
Tādējādi viens no organisma izmaiņu bioķīmiskajiem pamatiem treniņu ietekmē ir enzīmu sistēmu aktivitātes palielināšanās un darba laikā iztērēto enerģijas avotu superkompensācija. Kāpēc sporta treniņu praksē ir svarīgi ņemt vērā superkompensācijas modeļus?
Superkompensācijas modeļu pārzināšana ļauj zinātniski pamatot slodžu intensitāti un atpūtas intervālus normālu fizisko vingrinājumu un sporta treniņu laikā.
Tā kā superkompensācija saglabājas kādu laiku pēc darba beigām, turpmākos darbus var veikt labvēlīgākos bioķīmiskos apstākļos un, savukārt, novest pie tālākas funkcionālā līmeņa paaugstināšanās (.... att.). Ja turpmākais darbs tiek veikts nepilnīgas atveseļošanās apstākļos, tas noved pie funkcionālā līmeņa pazemināšanās (att.....).
Treniņu ietekmē organismā notiek aktīva adaptācija, bet nevis darbam “vispārīgi”, bet konkrētiem tā veidiem. Pētot dažāda veida sporta aktivitātes, tika noteikts bioķīmiskās adaptācijas specifikas princips un tika izveidoti motoriskās aktivitātes īpašību bioķīmiskie pamati - ātrums, spēks, izturība. Un tas nozīmē zinātniski pamatotus ieteikumus mērķtiecīgai apmācības sistēmai.
Sniegsim tikai vienu piemēru. Atcerieties, ka pēc intensīvas liela ātruma slodzes (skriešanas) palielinās elpošana ("elpas trūkums"). Ar ko tas saistīts? Darba laikā (skrienot) skābekļa trūkuma dēļ asinīs uzkrājas nepietiekami oksidēti produkti (pienskābe u.c.), kā arī oglekļa dioksīds, kas izraisa asins skābuma pakāpes izmaiņas. Attiecīgi tas izraisa iegarenās smadzenes elpošanas centra ierosmi un pastiprinātu elpošanu. Intensīvas oksidācijas rezultātā normalizējas asiņu skābums. Un tas ir iespējams tikai ar augstu aerobās oksidācijas enzīmu aktivitāti. Līdz ar to, beidzoties intensīvam darbam atpūtas periodā, aktīvi darbojas aerobās oksidācijas enzīmi. Tajā pašā laikā sportistu izturība, kas veic ilgstošu darbu, ir tieši atkarīga no aerobās oksidācijas aktivitātes. Pamatojoties uz to, tieši bioķīmiķi ieteica daudzu sporta veidu treniņos iekļaut īslaicīgas augstas intensitātes slodzes, kas šobrīd ir vispārpieņemts.
Kāda ir trenēta organisma bioķīmiskā īpašība?
Trenēta organisma muskuļos:
Palielinās miozīna saturs, brīvo HS-grupu skaits tajā, t.i. muskuļu spēja sadalīt ATP;
Palielinās ATP resintēzei nepieciešamo enerģijas avotu rezerves (kreatīna fosfāta, glikogēna, lipīdu u.c. saturs)
Ievērojami paaugstina enzīmu aktivitāti, kas katalizē gan anaerobos, gan aerobos oksidācijas procesus;
Muskuļos palielinās mioglobīna saturs, kas rada skābekļa rezerves muskuļos.
Muskuļu stromā palielinās olbaltumvielu saturs, kas nodrošina muskuļu relaksācijas mehānismu. Novērojumi sportistiem liecina, ka palielinās spēja atslābināt muskuļus treniņu ietekmē.
Pielāgošanās vienam faktoram palielina izturību pret citiem faktoriem (piemēram, stresu utt.);
Mūsdienu sportista treniņos ir nepieciešama augsta fizisko aktivitāšu intensitāte un liels to apjoms, kas var vienpusēji ietekmēt organismu. Tāpēc tas prasa pastāvīgu uzraudzību no ārstu, sporta medicīnas speciālistu puses, pamatojoties uz sporta bioķīmiju un fizioloģiju.
Un fiziskā izglītība, kā arī sporta aktivitātes ļauj attīstīt cilvēka ķermeņa rezerves spējas un nodrošināt viņam pilnīgu veselību, augstu veiktspēju un ilgmūžību. Fiziskā veselība ir neatņemama cilvēka personības harmoniskas attīstības sastāvdaļa, tā veido raksturu, garīgo procesu stabilitāti, gribas īpašības u.c.
Fiziskās audzināšanas un fiziskās kultūras medicīniskās un pedagoģiskās kontroles zinātniskās sistēmas dibinātājs ir ievērojams pašmāju zinātnieks, izcils skolotājs, anatoms un ārsts Petrs Francevičs Lesgafts. Viņa teorija balstās uz cilvēka fiziskās un garīgās, morālās un estētiskās attīstības vienotības principu. Viņš uzskatīja, ka fiziskās audzināšanas teorija ir "bioloģijas zinātnes nozare".
Milzīga loma bioloģijas zinātņu sistēmā, kas apgūst nodarbību pamatus fiziskās kultūras un sporta jomā, pieder bioķīmijai.
Jau pagājušā gadsimta 40. gados Ļeņingradas zinātnieka Nikolaja Nikolajeviča Jakovļeva laboratorijā tika uzsākti mērķtiecīgi zinātniski pētījumi sporta bioķīmijas jomā. Tie ļāva noskaidrot organisma pielāgošanās dažāda veida muskuļu aktivitātēm būtību un specifiskās iezīmes, pamatot sporta treniņu principus, faktorus, kas ietekmē sportista sniegumu, noguruma stāvokli, pārtrenēšanos un daudz ko citu. citi. uc Nākotnē sporta bioķīmijas attīstība veidoja pamatu astronautu sagatavošanai kosmosa lidojumiem.
Kādus jautājumus risina sporta bioķīmija?
Sporta bioķīmija ir sporta fizioloģijas un sporta medicīnas pamats. Darba muskuļu bioķīmiskajos pētījumos ir konstatēts:
Bioķīmisko izmaiņu modeļi kā aktīva adaptācija palielinātai muskuļu aktivitātei;
Sporta treniņu principu pamatojums (atkārtojums, regularitāte, darba un atpūtas attiecība u.c.)
Motoriskās aktivitātes īpašību bioķīmiskās īpašības (ātrums, spēks, izturība)
Veidi, kā paātrināt sportista ķermeņa atveseļošanos un ne tikai. citi
Jautājumi un uzdevumi.
Kāpēc ātrgaitas slodzes iedarbojas uz ķermeni daudzpusīgāk?
Mēģiniet sniegt fizioloģisku un bioķīmisku pamatojumu Aristoteļa apgalvojumam "Nekas neizsmeļ un nesagrauj cilvēku kā ilgstoša fiziska neaktivitāte." Kāpēc tas ir tik aktuāls mūsdienu cilvēkam?
NOmuskuļu šķiedru struktūra un kontrakcijas.
Muskuļu kontrakcija dzīvā sistēmā ir mehāniski ķīmisks process. Mūsdienu zinātne to uzskata par vispilnīgāko bioloģiskās mobilitātes veidu. Bioloģiskie objekti “attīstīja” muskuļu šķiedru kontrakciju kā veidu, kā pārvietoties telpā (kas būtiski paplašināja viņu dzīves iespējas).
Pirms muskuļu kontrakcijas notiek sasprindzinājuma fāze, kas ir darba rezultāts, kas tiek veikts, ķīmisko enerģiju tieši un ar labu efektivitāti (30-50%) pārvēršot mehāniskajā enerģijā. Potenciālās enerģijas uzkrāšanās sasprindzinājuma fāzē ieved muskuļus iespējamās, bet vēl neapzinātās kontrakcijas stāvoklī.
Dzīvniekiem un cilvēkiem ir (un cilvēki uzskata, ka tie jau ir labi izpētīti) divi galvenie muskuļu veidi: svītraina un gluda. svītraini muskuļi vai kauliem piestiprināts skelets (izņemot sirds muskuļa svītrainās šķiedras, kas pēc sastāva atšķiras no skeleta muskuļiem). Gluda muskuļus atbalsta iekšējo orgānu un ādas audus un veido asinsvadu sieniņu, kā arī zarnu muskuļus.
Sporta bioķīmijā viņi mācās skeleta muskuļi, "īpaši atbildīgs" par sportiskajiem rezultātiem.
Muskulis (kā makro veidojums, kas pieder pie makro objekta) sastāv no indivīda muskuļu šķiedras(mikro veidojumi). Muskuļos to ir tūkstošiem, attiecīgi, muskuļu piepūle ir neatņemama vērtība, kas summē daudzu atsevišķu šķiedru kontrakcijas. Ir trīs veidu muskuļu šķiedras: balts strauji sarūk , starpposma Un sarkans lēnām sarūk. Šķiedru veidi atšķiras pēc to enerģijas piegādes mehānisma un tiek kontrolēti ar dažādiem motoriem neironiem. Muskuļu veidi atšķiras pēc šķiedru veidu attiecības.
Atsevišķa muskuļu šķiedra - pavedienu bezšūnu veidošanās - simpplasts. Simplasts “neizskatās pēc šūnas”: tai ir stipri izstiepta forma garumā no 0,1 līdz 2-3 cm, sartorius muskulī līdz 12 cm un biezums no 0,01 līdz 0,2 mm. Simplastu ieskauj apvalks - sarkolemma kuras virsmai ir piemēroti vairāku kustību nervu gali. Sarkolemma ir divslāņu lipoproteīnu membrāna (10 nm bieza), kas pastiprināta ar kolagēna šķiedru tīklu. Atslābinoties pēc kontrakcijas, tie atgriež simplastu tā sākotnējā formā (4. att.).
Rīsi. 4. Atdaliet muskuļu šķiedru.
Sarkolemmas membrānas ārējā virsmā vienmēr tiek uzturēts elektriskās membrānas potenciāls, pat miera stāvoklī tas ir 90-100 mV. Potenciāla klātbūtne ir nepieciešams nosacījums muskuļu šķiedru (piemēram, automašīnas akumulatora) pārvaldībai. Potenciāls tiek radīts, pateicoties aktīvai (tas nozīmē, ar enerģijas izmaksām - ATP) vielu pārnešanai caur membrānu un tās selektīvajai caurlaidībai (pēc principa - "ielaidīšu vai izlaidīšu, ko gribēšu"). Tāpēc simpplasta iekšpusē daži joni un molekulas uzkrājas lielākā koncentrācijā nekā ārpusē.
Sarkolemma ir labi caurlaidīga K + joniem - tie uzkrājas iekšpusē, un Na + joni tiek noņemti ārpusē. Attiecīgi Na + jonu koncentrācija starpšūnu šķidrumā ir lielāka nekā K + jonu koncentrācija simplastā. PH nobīde uz skābes pusi (piemēram, pienskābes veidošanās laikā) palielina sarkolemmas caurlaidību makromolekulārām vielām (taukskābēm, olbaltumvielām, polisaharīdiem), kuras parasti caur to neiziet. Zemas molekulmasas vielas (glikoze, pienskābes un pirovīnskābes, ketonvielas, aminoskābes, īsie peptīdi) viegli izkļūst (izkliedējas) caur membrānu.
Simplasta iekšējais saturs ir sarkoplazma- šī ir koloidāla proteīna struktūra (pēc konsistences atgādina želeju). Suspendētā stāvoklī tas satur glikogēna ieslēgumus, tauku pilienus, tajā ir “iegultas” dažādas subcelulāras daļiņas: kodoli, mitohondriji, miofibrillas, ribosomas un citi.
Kontrakcijas "mehānisms" simplasta iekšpusē - miofibrils. Tie ir plāni (Ø 1 - 2 mikroni) muskuļu pavedieni, gari - gandrīz vienādi ar muskuļu šķiedras garumu. Konstatēts, ka netrenētu muskuļu simplastos miofibrillas nav sakārtotas sakārtoti, pa simpplastu, bet gan ar izplatību un novirzēm, savukārt trenētos miofibrillas ir orientētas pa garenasi un arī grupējas saišķos, piemēram, virvēs. (Vērpjot mākslīgās un sintētiskās šķiedras, polimēra makromolekulas sākotnēji neatrodas strikti gar šķiedru un, tāpat kā sportisti, tiek “spītīgi trenētas” - pareizi orientētas - pa šķiedru asi, atkārtoti pārtinot: garās darbnīcas skatīt plkst. ZIV un Khimvolokno).
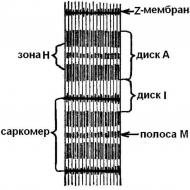
Gaismas mikroskopā var novērot, ka miofibrillas patiešām ir "šķērssvītras". Viņi mijas gaišās un tumšās zonas - diski. Tumši diski BET (anizotropās) olbaltumvielas satur vairāk nekā gaismas diskus es (izotropisks). Gaismas diski, ko šķērso membrānas Z (telofragmas) un miofibrilu laukums starp diviem Z - sauc par membrānām sarkomere. Miofibrils sastāv no 1000 - 1200 sarkomēriem (5. att.).
Muskuļu šķiedras kontrakcijas kopumā sastāv no vienas kontrakcijas sarkomēri. Savelkot katru atsevišķi, sarkomēri visi kopā rada neatņemamu spēku un veic mehānisku darbu, lai samazinātu muskuļus.
Sarkomēra garums svārstās no 1,8 µm miera stāvoklī līdz 1,5 µm ar mērenu kontrakciju un līdz 1 µm ar pilnīgu kontrakciju. Sarkomēru diski, tumši un gaiši, satur protofibrillas (miofilamentus) - proteīna pavedienveida struktūras. Tās ir divu veidu: biezas (Ø - 11 - 14 nm, garums - 1500 nm) un plānas (Ø - 4 - 6 nm, garums - 1000 nm).

Rīsi. 5. Miofibrilu parauglaukums.
gaismas diski ( es ) sastāv tikai no plānām protofibrilām un tumšiem diskiem ( BET ) - no divu veidu protofibrilām: plānas, kas savienotas kopā ar membrānu, un biezas, koncentrētas atsevišķā zonā ( H ).
Ar sarkomēra kontrakciju tumšā diska garums ( BET ) nemainās, kamēr gaismas diska garums ( es ) samazinās, plānām protofibrilām (gaišajiem diskiem) pārvietojoties spraugās starp biezajām (tumšajiem diskiem). Uz protofibrilu virsmas ir īpaši izaugumi - saaugumi (apmēram 3 nm biezumā). "Darba stāvoklī" tie veido saķeri (ar šķērseniskiem tiltiem) starp bieziem un plāniem protofibrilu pavedieniem (6. att.). Samazinot Z -membrānas balstās pret biezu protofibrilu galiem, un plānās protofibrillas var aptīties pat ap biezām. Pārmērīgas kontrakcijas laikā sarkomēra centrā esošo plānu pavedienu gali tiek ietīti, bet biezo protofibrilu gali tiek saburzīti.

Rīsi. 6. Smailu veidošanās starp aktīnu un miozīnu.
Muskuļu šķiedras tiek apgādātas ar enerģiju caur sarkoplazmatiskais tīkls(viņa arī ir sarkoplazmatiskais tīkls) - garenisko un šķērsenisko kanāliņu, membrānu, burbuļu, nodalījumu sistēmas.
Sarkoplazmatiskajā retikulā dažādi bioķīmiskie procesi norisinās organizēti un kontrolēti, tīkls aptver visu kopā un katru miofibrilu atsevišķi. Tīkls ietver ribosomas, tās veic olbaltumvielu sintēzi, bet mitohondriji - "šūnu enerģijas stacijas" (saskaņā ar skolas mācību grāmatas definīciju). Patiesībā mitohondriji ir uzbūvētas starp miofibrilām, kas rada optimālus apstākļus muskuļu kontrakcijas procesa enerģijas piegādei. Konstatēts, ka trenētos muskuļos mitohondriju skaits ir lielāks nekā tādos pašos netrenētajos.
Muskuļu ķīmiskais sastāvs.
ūdens ar atstāj 70-80% no muskuļu masas.
Vāveres. Olbaltumvielas veido 17 līdz 21% no muskuļu masas: aptuveni 40% no visiem muskuļu proteīniem ir koncentrēti miofibrilās, 30% sarkoplazmā, 14% mitohondrijās, 15% sarkolemmā, pārējie kodolos un citās šūnu organellās.
Muskuļu audi satur fermentus miogēnie proteīni grupas, mioalbumīns- uzglabāšanas proteīns (tā saturs pakāpeniski samazinās līdz ar vecumu), sarkanais proteīns mioglobīns- hromoproteīns (to sauc par muskuļu hemoglobīnu, tas saista skābekli vairāk nekā asins hemoglobīns), kā arī globulīni, miofibrilārie proteīni. Vairāk nekā puse no miofibrilārajiem proteīniem ir miozīns, apmēram ceturtdaļa aktīns, pārējais - tropomiozīns, troponīns, α- un β-aktinīni, fermenti kreatīna fosfokināze, deamināze un citi. Muskuļu audos ir kodolenerģijasvāveres- nukleoproteīni, mitohondriju proteīni. Olbaltumvielās stroma, muskuļu audu pīšana, - galvenā daļa - kolagēns Un elastīns sarkolemmas, kā arī miostromīni (saistīti ar Z - membrānas).
Inšķīstošie slāpekļa savienojumi. Cilvēka skeleta muskuļi satur dažādus ūdenī šķīstošos slāpekļa savienojumus: ATP no 0,25 līdz 0,4 % kreatīna fosfāts (CrF)- no 0,4 līdz 1% (treniņa laikā tā daudzums palielinās), to sabrukšanas produkti - ADP, AMP, kreatīns. Turklāt muskuļi satur dipeptīdu karnozīns, apmēram 0,1 - 0,3%, iesaistīti muskuļu darbaspējas atjaunošanā noguruma laikā; karnitīns, atbildīgs par taukskābju pārnešanu cauri šūnu membrānām; aminoskābes, un starp tām dominē glutamīnskābe (vai tas neizskaidro mononātrija glutamāta lietošanu, izlasiet garšvielu sastāvu, lai ēdienam piešķirtu gaļas garšu); purīna bāzes, urīnviela un amonjaks. Skeleta muskuļos ir arī aptuveni 1,5% fosfatīdi, iesaistīts audu elpošanā.
Nesatur slāpekli savienojumiem. Muskuļos ir ogļhidrāti, glikogēns un tā vielmaiņas produkti, kā arī tauki, holesterīns, ketonķermeņi un minerālsāļi. Atkarībā no uztura un treniņu pakāpes glikogēna daudzums svārstās no 0,2 līdz 3%, savukārt treniņš palielina brīvā glikogēna masu. Izturības treniņu laikā muskuļos uzkrājas rezerves tauki. Ar olbaltumvielām saistītie tauki ir aptuveni 1%, un muskuļu šķiedru membrānas var saturēt līdz 0,2% holesterīna.
Minerālvielas. Muskuļu audu minerālvielas veido aptuveni 1 - 1,5% no muskuļa svara, galvenokārt tie ir kālija, nātrija, kalcija, magnija sāļi. Minerālu joniem, piemēram, K +, Na +, Mg 2+, Ca 2+, Cl -, HP0 4 ~ ir svarīga loma bioķīmiskajos procesos muskuļu kontrakcijas laikā (tie ir iekļauti "sporta" piedevās un minerālūdenī).
Muskuļu proteīnu bioķīmija.
Galvenais saraušanās proteīns muskuļos miozīns attiecas uz fibrilāriem proteīniem (molekulmasa ir aptuveni 470 000). Svarīga miozīna iezīme ir spēja veidot kompleksus ar ATP un ADP molekulām (kas ļauj "atņemt" enerģiju no ATP), un ar proteīnu - aktīnu (kas ļauj noturēt kontrakciju).
Miozīna molekulai ir negatīvs lādiņš, un tā īpaši mijiedarbojas ar Ca ++ un Mg ++ joniem. Miozīns Ca ++ jonu klātbūtnē paātrina ATP hidrolīzi un tādējādi uzrāda fermentatīvu Adenozīna trifosfāta aktivitāte:
miozīns-ATP+H2O → miozīns + ADP + H3PO4 + darbs(enerģija 40 kJ/mol)
Miozīna proteīnu veido divas identiskas, garas polipeptīda α-ķēdes, kas savītas kā dubultspirāle, 7. att. Proteolītisko enzīmu ietekmē miozīna molekula sadalās divās daļās. Viena no tā daļām ar adhēziju palīdzību spēj saistīties ar aktīnu, veidojot aktomiozīnu. Šī daļa ir atbildīga par adenozīna trifosfatāzes aktivitāti, kas ir atkarīga no barotnes pH, optimālais ir pH 6,0 - 9,5, kā arī KCl koncentrācija. Akomiozīna komplekss sadalās ATP klātbūtnē, bet ir stabils, ja nav brīva ATP. Otrā miozīna molekulas daļa sastāv arī no divām savītām spirālēm, elektrostatiskā lādiņa dēļ tās saista miozīna molekulas protofibrilās.

Rīsi. 7. Akomiozīna struktūra.
Otrais lielākais saraušanās proteīns aktīns(7. att.). Tas var pastāvēt trīs veidos: monomērs (lodveida), dimērs (lodveida) un polimērs (fibrilārs). Monomērs globulārais aktīns, kad tā polipeptīdu ķēdes ir cieši iesaiņotas kompaktā sfēriskā struktūrā, ir saistīts ar ATP. Sašķeļ ATP, aktīna monomēri - A, veido dimērus, tai skaitā ADP: A - ADP - A. Polimēriskais fibrilārais aktīns ir dubultspirāle, kas sastāv no dimēriem, att. 7.
Lodveida aktīns K +, Mg ++ jonu klātbūtnē pārvēršas par fibrilāru aktīnu, dzīvajos muskuļos dominē fibrilārais aktīns.
Miofibrils satur ievērojamu daudzumu olbaltumvielu tropomiozīns, kas sastāv no divām - α-spirālveida polipeptīdu ķēdēm. Atpūtas muskuļos tas veido kompleksu ar aktīnu un bloķē tā aktīvos centrus, jo aktīns spēj saistīties ar Ca ++ joniem un tie noņem šo blokādi.
Molekulārā līmenī sarkomēra biezās un plānās protofibrillas mijiedarbojas elektrostatiski, jo tām ir īpašas zonas - izaugumi un izvirzījumi, kur veidojas lādiņš. A-diska zonā no gareniski orientētu miozīna molekulu kūlīša tiek veidotas biezas protofibrillas, ap biezām radiāli atrodas plānas protofibrillas, veidojot daudzkodolu kabelim līdzīgu struktūru. Resno protofibrilu centrālajā M joslā miozīna molekulas ir savienotas ar to "astes" palīdzību, un to izvirzītās "galvas" - izaugumi ir vērsti dažādos virzienos un atrodas pa regulārām spirālveida līnijām. Faktiski tiem pretī fibrilārā aktīna spirālēs noteiktā attālumā viena no otras izvirzās arī monomēra aktīna globulas. Katrai prezentācijai ir aktīvais centrs, kuru dēļ iespējama saauguma veidošanās ar miozīnu. Sarkomēru Z veida membrānas (piemēram, mainīgi pjedestāli) sastiprina kopā plānas protofibrillas.
Kontrakcijas un relaksācijas bioķīmija.
Cikliskās bioķīmiskās reakcijas, kas notiek muskulī kontrakcijas laikā, nodrošina atkārtotu saaugumu veidošanos un iznīcināšanu starp "galvām" - biezu protofibrilu miozīna molekulu izaugumiem un izvirzījumiem - tievo protofibrilu aktīviem centriem. Darbs pie adhēziju veidošanās un aktīna pavediena veicināšanas gar miozīna pavedienu prasa gan precīzu kontroli, gan ievērojamus enerģijas izdevumus. Reāli šķiedru saraušanās brīdī katrā aktīvajā centrā - dzega - veidojas aptuveni 300 saaugumi minūtē.
Kā jau minēts iepriekš, tikai ATP enerģiju var tieši pārvērst muskuļu kontrakcijas mehāniskajā darbā. ATP, ko hidrolizē miozīna enzīmu centrs, veido kompleksu ar visu proteīna miozīnu. ATP-miozīna kompleksā, kas piesātināts ar enerģiju, miozīns maina savu struktūru un līdz ar to ārējos “izmērus” un tādā veidā veic mehānisku darbu, lai saīsinātu miozīna pavediena izaugumu.
Atpūtas muskuļos miozīns joprojām ir saistīts ar ATP, bet caur Mg ++ joniem bez ATP hidrolītiskas šķelšanās. Miozīna un aktīna saķeres veidošanos miera stāvoklī novērš tropomiozīna komplekss ar troponīnu, kas bloķē aktīna aktīvos centrus. Blokāde tiek saglabāta un ATP netiek sadalīts, kamēr Ca ++ joni ir saistīti. Kad nervu impulss nonāk pie muskuļu šķiedras, tas atbrīvo impulsa raidītājs- neirohormons acetilholīns. Ar Na + joniem negatīvais lādiņš uz sarkolemmas iekšējās virsmas tiek neitralizēts un notiek tā depolarizācija. Šajā gadījumā Ca ++ joni atbrīvojas un saistās ar troponīnu. Savukārt troponīns zaudē lādiņu, kādēļ tiek atbrīvoti aktīvie centri - aktīna pavedienu izvirzījumi un starp aktīnu un miozīnu parādās saaugumi (jo tievo un biezo protofibrilu elektrostatiskā atgrūšana jau ir noņemta). Tagad Ca ++ klātbūtnē ATP mijiedarbojas ar miozīna fermentatīvās aktivitātes centru un sadalās, un pārveidotā kompleksa enerģija tiek izmantota, lai samazinātu saķeres. Iepriekš aprakstītā molekulāro notikumu ķēde ir līdzīga elektriskajai strāvai, kas uzlādē mikrokondensatoru, tās elektriskā enerģija uzreiz tiek pārvērsta mehāniskā darbā uz vietas un ir nepieciešams atkārtoti uzlādēt (ja vēlaties doties tālāk).
Pēc adhēzijas pārrāvuma ATP nesadalās, bet atkal veido enzīmu-substrāta kompleksu ar miozīnu:
M–A + ATP -----> M – ATP + A vai
M-ADP-A + ATP ----> M-ATP + A + ADP
Ja šajā brīdī pienāk jauns nervu impulss, tad “uzlādes” reakcijas atkārtojas, ja nākamais impulss nepienāk, muskulis atslābst. Savilktā muskuļa atgriešanos relaksācijas laikā sākotnējā stāvoklī nodrošina muskuļu stromas proteīnu elastīgie spēki. Izvirzot mūsdienu hipotēzes par muskuļu kontrakciju, zinātnieki ierosina, ka kontrakcijas brīdī aktīna pavedieni slīd pa miozīna pavedieniem, un to saīsināšana iespējama arī kontraktilo proteīnu telpiskās struktūras izmaiņu dēļ (spirāles formas izmaiņas).
Miera stāvoklī ATP ir plastificējoša iedarbība: savienojoties ar miozīnu, tas novērš tā adhēzijas veidošanos ar aktīnu. Sadaloties muskuļu kontrakcijas laikā, ATP nodrošina enerģiju adhēzijas saīsināšanas procesam, kā arī "kalcija sūkņa" darbam - Ca ++ jonu padevei. ATP šķelšanās muskuļos notiek ļoti lielā ātrumā: līdz 10 mikromoliem uz 1 g muskuļa minūtē. Tā kā kopējās ATP rezerves muskuļos ir nelielas (ar tām var pietikt tikai 0,5-1 sek darbam ar maksimālo jaudu), lai nodrošinātu normālu muskuļu darbību, ATP ir jāatjauno tādā pašā ātrumā, kādā tas tiek sadalīts.
Mācību grāmatā ir izklāstīti vispārējās bioķīmijas un cilvēka ķermeņa muskuļu darbības bioķīmijas pamati, aprakstīta svarīgāko organisma vielu ķīmiskā uzbūve un vielmaiņas procesi, atklāta to nozīme muskuļu darbības nodrošināšanā. Tiek aplūkoti muskuļu kontrakcijas procesu bioķīmiskie aspekti un enerģijas ģenerēšanas mehānismi muskuļos, motorisko īpašību attīstības modeļi, noguruma, atveseļošanās, adaptācijas procesi, kā arī racionāla uztura un sportistu funkcionālā stāvokļa diagnostika. apsvērts. Fiziskās audzināšanas un sporta augstākās un vidējās izglītības iestāžu studentiem un skolotājiem, fiziskās rehabilitācijas un atpūtas speciālistiem.
Grāmatas informācija:
Volkovs N.I., Nesens E.N., Osipenko A.A., Korsuns S.N. Muskuļu aktivitātes bioķīmija. 2000. - 503 lpp.
Pirmā daļa. Cilvēka ķermeņa dzīvībai svarīgās aktivitātes bioķīmiskie pamati
1. nodaļa. Ievads bioķīmijā
1. Bioķīmijas pētījuma priekšmets un metodes
2. Bioķīmijas attīstības vēsture un sporta bioķīmijas veidošanās
3. Cilvēka ķermeņa ķīmiskā uzbūve
4. Makromolekulu transformācija
testa jautājumi
2. nodaļa
1. Metabolisms ir dzīvā organisma pastāvēšanas nepieciešams nosacījums
2. Kataboliskās un anaboliskās reakcijas – vielmaiņas divas puses
3. Vielmaiņas veidi
4. Barības vielu sadalīšanās un enerģijas ieguves stadijas šūnās
5. Šūnu struktūras un to nozīme vielmaiņā
6. Metabolisma regulēšana
testa jautājumi
3. nodaļa
1. Enerģijas avoti
2. ATP – universāls enerģijas avots organismā
3. Bioloģiskā oksidēšana - galvenais enerģijas ražošanas veids ķermeņa šūnās
4. Mitohondriji - šūnas "enerģijas stacijas".
5. Citronskābes cikls ir galvenais barības vielu aerobās oksidācijas ceļš
6. Elpošanas ķēde
7. Oksidatīvā fosforilēšana ir galvenais ATP sintēzes mehānisms
8. ATP metabolisma regulēšana
testa jautājumi
4. nodaļa
1. Ūdens un tā loma organismā
2. Ūdens bilance un tā maiņa muskuļu darbības laikā
3. Minerālvielas un to nozīme organismā
4. Minerālu vielmaiņa muskuļu darbības laikā
testa jautājumi
5. nodaļa
1. Vielu transportēšanas mehānismi
2. Ķermeņa iekšējās vides skābju-bāzes stāvoklis
3. Bufersistēmas un to loma barotnes nemainīga pH uzturēšanā
testa jautājumi
6. nodaļa
1. Vispārīga izpratne par fermentiem
2. Fermentu un koenzīmu struktūra
3. Vairākas enzīmu formas
4. Fermentu īpašības
5. Fermentu darbības mehānisms
6. Fermentu darbību ietekmējošie faktori
7. Fermentu klasifikācija
testa jautājumi
7. nodaļa
1. Vispārīga izpratne par vitamīniem
2. Vitamīnu klasifikācija
3. Taukos šķīstošo vitamīnu raksturojums
4. Ūdenī šķīstošo vitamīnu raksturojums
5. Vitamīniem līdzīgas vielas
testa jautājumi
8. nodaļa
1. Izpratne par hormoniem
2. Hormonu īpašības
3. Hormonu ķīmiskā daba
4. Hormonu biosintēzes regulēšana
5. Hormonu darbības mehānisms
6. Hormonu bioloģiskā loma
7. Hormonu loma muskuļu darbībā
testa jautājumi
9. nodaļa
1. Ogļhidrātu ķīmiskais sastāvs un bioloģiskā loma
2. Ogļhidrātu klašu raksturojums
3. Ogļhidrātu vielmaiņa cilvēka organismā
4. Ogļhidrātu sadalīšanās gremošanas laikā un to uzsūkšanās asinīs
5. Glikozes līmenis asinīs un tā regulēšana
6. Ogļhidrātu intracelulārais metabolisms
7. Ogļhidrātu vielmaiņa muskuļu aktivitātes laikā
testa jautājumi
10. nodaļa
1. Lipīdu ķīmiskais sastāvs un bioloģiskā loma
2. Lipīdu klašu raksturojums
3. Tauku vielmaiņa organismā
4. Tauku sadalīšanās gremošanas laikā un to uzsūkšanās
5. Intracelulārā tauku vielmaiņa
6. Lipīdu metabolisma regulēšana
7. Lipīdu metabolisma pārkāpums
8. Tauku vielmaiņa muskuļu darbības laikā
testa jautājumi
11. nodaļa
1. Nukleīnskābju ķīmiskā struktūra
2. DNS struktūra, īpašības un bioloģiskā loma
3. RNS struktūra, īpašības un bioloģiskā loma
4. Nukleīnskābju apmaiņa
testa jautājumi
12. nodaļa
1. Olbaltumvielu ķīmiskais sastāvs un bioloģiskā loma
2. Aminoskābes
3. Olbaltumvielu strukturālā organizācija
4. Olbaltumvielu īpašības
5. Atsevišķu muskuļu darbā iesaistīto proteīnu raksturojums
6. Brīvie peptīdi un to nozīme organismā
7. Olbaltumvielu metabolisms organismā
8. Olbaltumvielu sadalīšanās gremošanas un aminoskābju uzsūkšanās laikā
9. Olbaltumvielu biosintēze un tās regulēšana
10. Intersticiāla proteīna sadalīšanās
11. Aminoskābju intracelulāra konversija un urīnvielas sintēze
12. Olbaltumvielu metabolisms muskuļu darbības laikā
testa jautājumi
13. nodaļa. Metabolisma integrācija un regulēšana - adaptācijas procesu bioķīmiskais pamats
1. Ogļhidrātu, tauku un olbaltumvielu savstarpēja konversija
2. Vielmaiņas regulējošās sistēmas un to nozīme organisma pielāgošanā fiziskajam stresam
3. Atsevišķu audu loma starpproduktu metabolisma integrācijā
testa jautājumi
Otrā daļa. Sporta bioķīmija
14. nodaļa
1. Muskuļu un muskuļu šķiedru veidi
2. Muskuļu šķiedru strukturālā organizācija
3. Muskuļu audu ķīmiskais sastāvs
4. Strukturālās un bioķīmiskās izmaiņas muskuļos kontrakcijas un relaksācijas laikā
5. Muskuļu kontrakcijas molekulārais mehānisms
testa jautājumi
15. nodaļa
1. Enerģijas ražošanas mehānismu vispārīgie raksturojumi
2. ATP resintēzes kreatīnfosfokināzes mehānisms
3. ATP resintēzes glikolītiskais mehānisms
4. ATP resintēzes miokināzes mehānisms
5. ATP resintēzes aerobais mehānisms
6. Enerģētisko sistēmu pieslēgšana dažādu fizisko slodžu laikā un to pielāgošana treniņu laikā
testa jautājumi
16. nodaļa
1. Bioķīmisko procesu izmaiņu vispārējais virziens muskuļu darbības laikā
2. Skābekļa transportēšana uz strādājošiem muskuļiem un tā patēriņš muskuļu darbības laikā
3. Bioķīmiskās izmaiņas atsevišķos orgānos un audos muskuļu darba laikā
4. Fizisko vingrinājumu klasifikācija pēc bioķīmisko izmaiņu rakstura muskuļu darba laikā
testa jautājumi
17. nodaļa
1. Noguruma bioķīmiskie faktori īstermiņa maksimālās un submaksimālās jaudas vingrinājumu laikā
2. Noguruma bioķīmiskie faktori ilgstošas lielas un mērenas jaudas vingrinājumu laikā
testa jautājumi
18. nodaļa
1. Bioķīmisko atveseļošanās procesu dinamika pēc muskuļu darba
2. Enerģijas rezervju atjaunošanas secība pēc muskuļu darba
3. Sabrukšanas produktu likvidēšana atpūtas periodā pēc muskuļu darba
4. Atveseļošanās procesu plūsmas iezīmju izmantošana sporta treniņu konstruēšanā
testa jautājumi
19. nodaļa
1. Personas fizisko veiktspēju ierobežojošie faktori
2. Sportista aerobā un anaerobā snieguma rādītāji
3. Treniņu ietekme uz sportistu sniegumu
4. Vecums un sportiskais sniegums
testa jautājumi
20. nodaļa
1. Ātruma-stiprības īpašību bioķīmiskās īpašības
2. Sportistu ātruma-spēka treniņu metožu bioķīmiskie pamati
testa jautājumi
21. nodaļa
1. Bioķīmiskie izturības faktori
2. Treniņu metodes, kas veicina izturību
testa jautājumi
22. nodaļa
1. Fiziskās aktivitātes, adaptācijas un treniņu efekts
2. Bioķīmiskās adaptācijas attīstības modeļi un apmācības principi
3. Adaptīvo izmaiņu specifika organismā treniņa laikā
4. Adaptīvo izmaiņu atgriezeniskums treniņa laikā
5. Adaptīvo izmaiņu secība treniņa laikā
6. Treniņu efektu mijiedarbība treniņa laikā
7. Adaptācijas cikliskā attīstība apmācības procesā
testa jautājumi
23. nodaļa
1. Sportistu racionāla uztura principi
2. Organisma enerģijas patēriņš un tā atkarība no veiktā darba
3. Uzturvielu līdzsvars sportista uzturā
4. Atsevišķu pārtikas ķīmisko komponentu loma muskuļu aktivitātes nodrošināšanā
5. Uztura bagātinātāji un svara kontrole
testa jautājumi
24. nodaļa
1. Bioķīmiskās kontroles uzdevumi, veidi un organizācija
2. Pētījuma objekti un galvenie bioķīmiskie parametri
3. Galvenie asins un urīna sastāva bioķīmiskie rādītāji, to izmaiņas muskuļu darbības laikā
4. Organisma energoapgādes sistēmu attīstības bioķīmiskā kontrole muskuļu darbības laikā
5. Bioķīmiskā kontrole pār treniņu līmeni, sportista organisma nogurumu un atjaunošanos
6. Dopinga kontrole sportā
testa jautājumi
Terminu vārdnīca
Vienības
Literatūra
Vairāk par grāmatu: formāts: pdf, faila izmērs: 37,13 Mb.
Saistītie raksti