講義:スポーツトレーニングの生化学的基礎。 筋肉の働きの間の体内の生化学的プロセスのダイナミクス筋肉の働きの生化学
アスリートの体はどのように激しい筋活動に適応しますか?
増加した筋肉活動に適応する過程で生じた身体の深い機能的変化は、スポーツの生理学によって研究されています。 しかし、それらは組織や臓器、そして最終的には体全体の代謝における生化学的変化に基づいています。 ただし、最も一般的な形式では、筋肉のみのトレーニングの影響下で発生する主な変化を検討します。
トレーニングの影響下での筋肉の生化学的再構築は、支出のプロセスと筋肉の機能的およびエネルギーの蓄えの回復の相互依存性に基づいています。 前の記事からすでに理解しているように、筋活動中にATPの集中的な分裂が起こり、それに応じて他の物質が集中的に消費されます。 筋肉では、クレアチンリン酸、グリコーゲン、脂質です。肝臓では、グリコーゲンが分解されて糖を形成し、血液とともに骨格筋、心臓、脳に移動します。 脂肪は分解され、脂肪酸は酸化されます。 同時に、代謝産物が体内に蓄積します-リン酸と乳酸、ケトン体、二酸化炭素。 一部は体によって失われ、一部は再び使用され、代謝に関与します。 筋肉の活動は多くの酵素の活動の増加を伴い、これにより使用済み物質の合成が始まります。 ATP、クレアチンリン酸、グリコーゲンの再合成は作業中にすでに可能ですが、これに伴い、これらの物質の集中的な分解があります。 したがって、作業中の筋肉の内容が元の内容に達することはありません。
残りの期間、エネルギー源の集中的な分割が停止すると、再合成のプロセスが明確な優位性を獲得し、消費されたものの回復(補償)が発生するだけでなく、初期を超える超回復(超補償)も発生しますレベル。 このパターンは「超回復の法則」と呼ばれます。
超回復の現象の本質。
スポーツの生化学では、このプロセスの規則性が研究されてきました。 たとえば、筋肉、肝臓、および他の臓器に物質が集中的に消費される場合、再合成がより速く進行し、過剰回復の現象がより顕著になることが確立されています。 たとえば、短期間の集中的な作業の後、筋肉内のグリコーゲンのレベルが最初のレベルを超えて増加し、1時間の休息後に発生し、12時間後に最初の最終レベルに戻ります。 長時間働いた後、超回復は12時間後にのみ発生しますが、筋肉内のグリコーゲンレベルの上昇は3日以上持続します。 これは、酵素の高い活性とそれらの強化された合成のためにのみ可能です。
したがって、トレーニングの影響下での身体の変化の生化学的基盤の1つは、酵素システムの活性の増加と、作業中に消費されるエネルギー源の超補償です。 スポーツトレーニングの実践において、超回復のパターンを考慮することが重要なのはなぜですか?
超回復のパターンを知ることで、通常の運動中やスポーツトレーニング中の負荷の強さと休息の間隔を科学的に実証することができます。
超回復は作業終了後もしばらく続くため、その後の作業はより好ましい生化学的条件で行うことができ、機能レベルのさらなる向上につながります(図...)。 回復が不完全な状態でその後の作業を行うと、機能レベルが低下します(図...)。
トレーニングの影響下で、身体の中で積極的な適応が起こりますが、「一般的に」機能するのではなく、特定の種類の適応に機能します。 さまざまな種類のスポーツ活動を研究する際に、生化学的適応の特異性の原則が確立され、運動活動の質の生化学的基盤が確立されました-速度、強さ、持久力。 そしてそれは、対象を絞ったトレーニングシステムに対する科学に基づく推奨を意味します。
一例を挙げましょう。 激しい高速負荷(ランニング)の後、呼吸が増加する(「息切れ」)ことを覚えておいてください。 それは何と関係がありますか? 作業(ランニング)中は、血液中の酸素不足により、酸素不足の生成物(乳酸など)や二酸化炭素が蓄積し、血液の酸性度が変化します。 したがって、これは延髄の呼吸中枢の興奮と呼吸の増加を引き起こします。 激しい酸化の結果として、血液の酸性度は正常化されます。 そして、これは好気性酸化酵素の高い活性でのみ可能です。 その結果、休息期間中の集中的な作業の終わりに、好気性酸化の酵素が活発に機能しています。 同時に、長期的な仕事をするアスリートの耐久性は、好気性酸化の活動に直接依存します。 これに基づいて、現在一般的に受け入れられている多くのスポーツのトレーニングに短期間の高強度の負荷を含めることを推奨したのは生化学者でした。
訓練された生物の生化学的特徴は何ですか?
訓練された生物の筋肉:
ミオシンの含有量、その中の遊離HSグループの数、すなわち ATPを分割する筋肉の能力;
ATP再合成に必要なエネルギー源の蓄えが増加します(クレアチンリン酸、グリコーゲン、脂質などの含有量)
嫌気性と好気性の両方の酸化プロセスを触媒する酵素の活性を大幅に高めます。
筋肉内のミオグロビンの含有量が増加し、筋肉内に酸素が蓄えられます。
筋弛緩のメカニズムを提供する筋ストロマのタンパク質の含有量が増加します。 アスリートの観察によると、トレーニングの影響下で筋肉をリラックスさせる能力が向上します。
ある要因に適応すると、他の要因(たとえば、ストレスなど)に対する抵抗力が高まります。
現代のアスリートのトレーニングには、高強度の身体活動と大量の運動が必要であり、それは体に一方的な影響を与える可能性があります。 したがって、スポーツの生化学と生理学に基づいて、医師、スポーツ医学の専門家による継続的な監視が必要です。
そして、体育やスポーツ活動は、人体の予備力を発達させ、彼に完全な健康、高性能、そして長寿を提供することを可能にします。 身体的健康は人格の調和のとれた発達の不可欠な部分であり、それは性格、精神的プロセスの安定性、意欲的な資質などを形成します。
体育の科学システムと、体育における医学的および教育学的管理の創設者は、注目に値する国内の科学者であり、優れた教師、解剖学者、および医師であるPetrFrantsevichLesgaftです。 彼の理論は、人の肉体的、精神的、道徳的、美的発達の統一の原則に基づいています。 彼は体育の理論を「生物科学の分派」と見なした。
物理文化やスポーツの分野で授業の基礎を研究する生物科学のシステムにおける大きな役割は、生化学に属しています。
すでに前世紀の40年代に、スポーツ生化学の分野での意図的な科学研究が、レニングラードの科学者ニコライニコライエビッチヤコブレフの研究室で開始されました。 彼らは、さまざまな種類の筋肉活動への身体の適応の本質と特定の特徴を見つけ出し、スポーツトレーニングの原則、アスリートのパフォーマンスに影響を与える要因、疲労状態、オーバートレーニングなどを実証することを可能にしましたその他。 将来的には、スポーツの生化学の発展は、宇宙飛行士の宇宙飛行の準備の基礎を形成しました。
スポーツの生化学はどのような質問を解決しますか?
スポーツ生化学は、スポーツ生理学とスポーツ医学の基礎です。 骨格筋の生化学的研究では、以下が確立されています。
増加した筋活動への積極的な適応としての生化学的変化のパターン。
スポーツトレーニングの原則の実証(繰り返し、規則性、仕事と休息の比率など)
運動活動の質(速度、強さ、持久力)の生化学的特性
アスリートの体の回復などを加速する方法。 その他
質問とタスク。
なぜ高速負荷がより用途の広い体に作用するのですか?
アリストテレスの「長時間の運動不足のように人を疲れさせたり破壊したりするものはない」という声明を生理学的および生化学的正当化するようにしてください。 なぜそれは現代人にそれほど関連しているのですか?
から筋線維の構造と収縮。
生体系の筋収縮は機械化学的プロセスです。 現代科学は、それを生物学的移動性の最も完璧な形と見なしています。 生物学的オブジェクトは、宇宙を移動する方法として筋線維の収縮を「発達」させました(これにより、生命の可能性が大幅に拡大しました)。
筋収縮の前に張力段階があります。これは、化学エネルギーを機械エネルギーに直接かつ効率的に(30〜50%)変換することによって実行される作業の結果です。 緊張期に位置エネルギーが蓄積すると、筋肉は収縮する可能性がありますが、まだ実現されていません。
動物と人間は持っています(そして人間は彼らがすでによく研究されていると信じています) 筋肉の2つの主なタイプ:縞模様で滑らか。 横紋筋または骨に付着した骨格筋(骨格筋とは組成が異なる心筋の横紋筋を除く)。 スムーズ 筋肉内臓や皮膚の組織を支え、血管壁や腸の筋肉を形成します。
スポーツの生化学では、彼らは研究します 骨格筋、スポーツの結果に対して「特に責任がある」。
筋肉(マクロオブジェクトに属するマクロフォーメーションとして)は、個人で構成されます 筋線維(マイクロフォーメーション)。 筋肉にはそれぞれ何千ものそれらがあり、筋肉の努力は多くの個々の繊維の収縮を合計する不可欠な値です。 筋線維には3つのタイプがあります: 白急速に減少 , 中級と 赤ゆっくりと縮みます。 繊維の種類は、エネルギー供給のメカニズムが異なり、さまざまな運動ニューロンによって制御されます。 筋肉の種類は、繊維の種類の比率が異なります。
分離した筋線維-糸状の無細胞形成- シンプラスト。 シンプラストは「細胞のようには見えない」:長さ0.1〜2〜3cm、縫工筋で最大12cm、厚さ0.01〜0.2mmの非常に細長い形状をしている。 シンプラストはシェルに囲まれています- 筋鞘いくつかの運動神経の終末が適している表面に。 筋鞘は、コラーゲン繊維のネットワークで強化された二重層リポタンパク質膜(厚さ10 nm)です。 収縮後に弛緩すると、シンプラストは元の形状に戻ります(図4)。
米。 4.筋線維を分離します。
筋鞘膜の外面では、電気膜電位は常に維持されており、安静時でも90〜100mVです。 電位の存在は、(自動車のバッテリーのような)筋線維の管理に必要な条件です。 可能性は、膜を介した物質の能動的(つまりエネルギーのコスト-ATP)移動とその選択的透過性(原則に従って-「私は好きな人を出し入れする」)によって作成されます。 したがって、シンプラストの内部では、一部のイオンと分子が外部よりも高濃度で蓄積します。
筋鞘はK+イオンに対して十分に透過性があり、内部に蓄積し、Na+イオンは外部から除去されます。 したがって、細胞間液中のNa +イオンの濃度は、シンプラスト内のK+イオンの濃度よりも高くなります。 pHを酸側にシフトすると(たとえば、乳酸の形成中)、通常は筋鞘を通過しない高分子物質(脂肪酸、タンパク質、多糖類)の筋鞘の透過性が高まります。 低分子量の物質(グルコース、乳酸、ピルビン酸、ケトン体、アミノ酸、短いペプチド)は、膜を簡単に通過(拡散)します。
シンプラストの内部内容は 筋形質-これはコロイド状のタンパク質構造です(一貫性のあるゼリーに似ています)。 浮遊状態では、グリコーゲン封入体、脂肪滴が含まれ、核、ミトコンドリア、筋原線維、リボソームなどのさまざまな細胞内粒子が「埋め込まれ」ています。
シンプラスト内の収縮性「メカニズム」- 筋原線維。これらは細い(Ø1〜2ミクロン)筋フィラメントで、長い-筋線維の長さにほぼ等しい。 訓練されていない筋肉のシンプラストでは、筋原線維はシンプラストに沿って整然と配置されていないが、広がりと偏差があり、訓練されたものでは、筋原線維は縦軸に沿って配向され、ロープで。 (人工繊維と合成繊維を紡ぐ場合、ポリマー高分子は最初は繊維に沿って厳密に配置されておらず、アスリートのように、巻き戻しを繰り返すことにより、繊維の軸に沿って「頑固に訓練」され、正しく配向されます。 ZIVおよびKhimvolokno)。
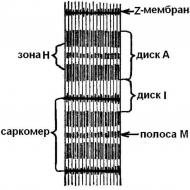
光学顕微鏡では、筋原線維が実際に「クロスストライプ」されていることがわかります。 それらは明るい領域と暗い領域を交互に繰り返します-ディスク。 暗いリム しかし (異方性)タンパク質には、ライトディスク以上のものが含まれています 私 (等方性)。 膜が交差するライトディスク Z (telophragms)と2つの間の筋原線維の領域 Z -膜と呼ばれる サルコメア。 筋原線維は1000〜1200のサルコメアで構成されています(図5)。
全体としての筋線維の収縮は、単一の収縮で構成されています サルコメア。それぞれを別々に収縮させると、サルコメアはすべて一緒に統合された力を生み出し、筋肉を減らすための機械的仕事を実行します。
サルコメアの長さは、安静時の1.8 µmから中程度の収縮の場合の1.5 µm、完全な収縮の場合の最大1 µmまで変化します。 サルコメアの円盤は、暗くても明るくても、プロトフィブリル(筋フィラメント)(タンパク質の糸状構造)を含んでいます。 それらには、厚い(Ø-11-14 nm、長さ-1500 nm)と薄い(Ø-4-6 nm、長さ-1000 nm)の2つのタイプがあります。

米。 5.筋原線維のプロット。
ライトディスク( 私 )薄いプロトフィブリルと暗いディスク( しかし ) -2つのタイプのプロトフィブリルから:薄い、膜で一緒に固定されている、および厚い、別のゾーンに集中している( H ).
サルコメア収縮では、暗い円盤の長さ( しかし ) ライトディスクの長さ( 私 )薄いプロトフィブリル(明るいディスク)が厚いプロトフィブリル(暗いディスク)の間のギャップに移動すると減少します。 プロトフィブリルの表面には、癒着(約3 nmの厚さ)という特別な成長が見られます。 「作業位置」では、それらはプロトフィブリルの太いフィラメントと細いフィラメントの間に(横方向のブリッジによって)係合を形成します(図6)。 減らすとき Z -膜は厚いプロトフィブリルの端に寄りかかっており、薄いプロトフィブリルは厚いプロトフィブリルを包み込むことさえできます。 過収縮時には、サルコメアの中央にある細いフィラメントの端が包まれ、太いプロトフィブリルの端がくしゃくしゃになります。

米。 6.アクチンとミオシンの間のスパイクの形成。
筋線維にはエネルギーが供給されます 筋小胞体(彼女も 筋小胞体)-縦行および横行小管、膜、気泡、コンパートメントのシステム。
筋小胞体では、さまざまな生化学的プロセスが組織化され制御された方法で進行し、ネットワークはすべてを一緒にカバーし、各筋原線維を別々にカバーします。 網状組織にはリボソームが含まれ、それらはタンパク質の合成を実行し、ミトコンドリア-「細胞エネルギーステーション」(学校の教科書の定義による)。 実際 ミトコンドリア筋原線維の間に構築され、筋収縮プロセスのエネルギー供給に最適な条件を作成します。 訓練された筋肉のミトコンドリアの数は、同じ訓練されていない筋肉のミトコンドリアの数よりも多いことが確立されています。
筋肉の化学組成。
と水筋肉重量の70-80%を残します。
リス。 タンパク質は筋肉重量の17〜21%を占めます。すべての筋肉タンパク質の約40%は筋原線維に、30%は筋形質に、14%はミトコンドリアに、15%は筋鞘に、残りは核やその他の細胞小器官に集中しています。
筋肉組織には酵素が含まれています 筋原性タンパク質グループ、 ミオアルブミン-貯蔵タンパク質(その含有量は年齢とともに徐々に減少します)、赤色タンパク質 ミオグロビン-色素タンパク質(筋肉ヘモグロビンと呼ばれ、血液ヘモグロビンよりも酸素と結合します)、および グロブリン、筋原線維タンパク質。筋原線維タンパク質の半分以上は ミオシン、約4分の1 アクチン、残り-トロポミオシン、トロポニン、α-およびβ-アクチニン、酵素 クレアチンホスホキナーゼ、デアミナーゼなど。 筋肉組織には 核リス-核タンパク質、 ミトコンドリアタンパク質。タンパク質では 間質、筋組織を編む-主要部分- コラーゲンと エラスチン筋鞘、およびミオストロミン( Z -膜)。
の可溶性窒素化合物。人間の骨格筋には、さまざまな水溶性窒素化合物が含まれています。 ATP 0.25から0.4%まで、 クレアチンリン酸(CrF)-0.4から1%(トレーニング中、その量は増加します)、それらの崩壊生成物-ADP、AMP、クレアチン。 さらに、筋肉にはジペプチドが含まれています カルノシン、約0.1〜0.3%、倦怠感時の筋肉のパフォーマンスの回復に関与します。 カルニチン、細胞膜を通過する脂肪酸の移動に関与します。 アミノ酸、そしてそれらの中でグルタミン酸が優勢です(これはグルタミン酸ナトリウムの使用を説明していません、食べ物に肉の味を与えるために調味料の組成を読んでください); プリン塩基、尿素、アンモニア。 骨格筋にも約1.5%含まれています ホスファチド、組織の呼吸に関与します。
窒素フリー 接続。 筋肉には、炭水化物、グリコーゲン、その代謝産物、脂肪、コレステロール、ケトン体、ミネラル塩が含まれています。 食事とトレーニングの程度に応じて、グリコーゲンの量は0.2から3%まで変化しますが、トレーニングは遊離グリコーゲンの量を増やします。 持久力トレーニング中に筋肉の予備脂肪が蓄積します。 タンパク質結合脂肪は約1%であり、筋線維膜には最大0.2%のコレステロールが含まれている可能性があります。
ミネラル。筋肉組織のミネラル物質は、筋肉の重量の約1〜1.5%を占め、これらは主にカリウム、ナトリウム、カルシウム、マグネシウムの塩です。 K +、Na +、Mg 2+、Ca 2+、Cl-、HP0 4〜などのミネラルイオンは、筋収縮中の生化学的プロセスで重要な役割を果たします(これらは「スポーツ」サプリメントとミネラルウォーターに含まれています)。
筋肉タンパク質の生化学。
筋肉の主要な収縮タンパク質 ミオシン繊維状タンパク質を指します(分子量は約470,000です)。 ミオシンの重要な特徴は、ATPおよびADP分子(ATPからエネルギーを「奪う」ことを可能にする)、およびタンパク質-アクチン(収縮を保持することを可能にする)と複合体を形成する能力です。
ミオシン分子は負の電荷を持ち、Ca++およびMg++イオンと特異的に相互作用します。 Ca ++イオンの存在下でのミオシンは、ATPの加水分解を促進し、したがって酵素を示します アデノシン三リン酸活性:
ミオシン-ATP+ H2O →ミオシン+ADP+ H3PO4 +仕事(エネルギー40 kJ / mol)
ミオシンタンパク質は、二重らせんのようにねじれた2つの同一の長いポリペプチドα鎖によって形成されます(図7)。 タンパク質分解酵素の作用下で、ミオシン分子は2つの部分に分解します。 その部分の1つは、癒着を介してアクチンに結合し、アクトミオシンを形成することができます。 この部分は、培地のpHに依存するアデノシントリホスファターゼ活性に関与し、最適値はpH 6.0〜9.5、およびKClの濃度です。 アクトミオシン複合体はATPの存在下で分解しますが、遊離ATPの非存在下では安定しています。 ミオシン分子の2番目の部分も、2つのねじれたらせんで構成されています。静電荷のため、ミオシン分子をプロトフィブリルに結合します。

米。 7.アクトミオシンの構造。
2番目の主要な収縮性タンパク質 アクチン(図7)。 それは、単量体(球状)、二量体(球状)、および高分子(繊維状)の3つの形態で存在する可能性があります。 単量体の球状アクチンは、そのポリペプチド鎖がコンパクトな球状構造にしっかりと詰め込まれている場合、ATPと関連しています。 分割ATP、アクチンモノマー-A、ADPを含むダイマーを形成します:A-ADP-A。高分子繊維状アクチンは、ダイマーからなる二重らせんです。 7。
球状アクチンは、K +、Mg ++イオンの存在下で繊維状アクチンに変化し、繊維状アクチンは生きている筋肉で優勢です。
筋原線維にはかなりの量のタンパク質が含まれています トロポミオシン、2つの-α-ヘリックスポリペプチド鎖で構成されています。 静止している筋肉では、アクチンと複合体を形成し、その活性中心をブロックします。これは、アクチンがCa ++イオンに結合でき、このブロックを取り除くためです。
分子レベルでは、サルコメアの太いプロトフィブリルと細いプロトフィブリルは、電荷が形成される特別な領域(成長と突起)を持っているため、静電的に相互作用します。 Aディスクの領域では、太いプロトフィブリルは縦方向に配向したミオシン分子の束から構築され、細いプロトフィブリルは太いものの周りに放射状に配置され、マルチコアケーブルに似た構造を形成します。 太いプロトフィブリルの中央のMバンドでは、ミオシン分子は「尾」と突き出た「頭」によって接続されています。副産物はさまざまな方向に向けられ、規則的ならせん状の線に沿って配置されています。 実際、繊維状アクチンのらせんの反対側で、互いに一定の距離を置いて、単量体のアクチン小球も突き出ています。 各プレゼンテーションには アクティブセンター、そのため、ミオシンとの癒着の形成が可能です。 サルコメアのZ膜(交互の台座のような)は、薄いプロトフィブリルを一緒に固定します。
収縮と弛緩の生化学。
収縮中に筋肉で発生する周期的な生化学反応は、「頭」(厚いプロトフィブリルのミオシン分子の成長と突起)の間の癒着の繰り返しの形成と破壊を確実にします-薄いプロトフィブリルの活性中心。 癒着の形成とミオシンフィラメントに沿ったアクチンフィラメントの促進に関する作業には、正確な制御とかなりのエネルギー消費の両方が必要です。 実際には、繊維の収縮の瞬間に、毎分約300の癒着が各アクティブセンター(レッジ)で形成されます。
先に述べたように、ATPのエネルギーだけが直接筋収縮の機械的仕事に変換することができます。 ミオシンの酵素中心によって加水分解されたATPは、タンパク質ミオシン全体と複合体を形成します。 エネルギーで飽和したATP-ミオシン複合体では、ミオシンはその構造を変化させ、それによって外部の「寸法」を変化させ、このようにして、ミオシンフィラメントの伸長を短縮するための機械的作業を実行します。
安静時の筋肉では、ミオシンは依然としてATPと関連していますが、ATPの加水分解による切断なしにMg++イオンを介しています。 安静時のミオシンとアクチンの間の癒着の形成は、トロポミオシンとアクチンの活性中心を遮断するトロポニンとの複合体によって防止されます。 Ca ++イオンが結合している間、封鎖は維持され、ATPは分割されません。 神経インパルスが筋線維に到達すると、それは解放されます パルストランスミッター–神経ホルモン アセチルコリン。 Na +イオンを使用すると、筋鞘の内面の負電荷が中和され、その脱分極が発生します。 この場合、Ca ++イオンが放出され、トロポニンに結合します。 次に、トロポニンはその電荷を失います。これが、活性中心-アクチンフィラメントの突起が解放され、アクチンとミオシンの間に癒着が現れる理由です(薄くて厚いプロトフィブリルの静電反発力はすでに除去されているため)。 現在、Ca ++の存在下で、ATPはミオシン酵素活性の中心と相互作用して分裂し、変換された複合体のエネルギーが癒着を減らすために使用されます。 上記の一連の分子イベントは、マイクロコンデンサーを再充電する電流に似ており、その電気エネルギーはその場ですぐに機械的仕事に変換され、再度充電する必要があります(先に進みたい場合)。
接着が破れた後、ATPは分裂しませんが、ミオシンと酵素-基質複合体を再び形成します。
M–A + ATP -----> M – ATP + Aまた
M-ADP-A + ATP ----> M-ATP + A + ADP
この時点で新しい神経インパルスが到着すると、「再充電」反応が繰り返され、次のインパルスが到着しない場合、筋肉が弛緩します。 弛緩中の収縮した筋肉の元の状態への復帰は、筋間質のタンパク質の弾性力によって提供されます。 筋収縮の現代の仮説を提唱し、科学者は、収縮の瞬間に、アクチンフィラメントがミオシンフィラメントに沿ってスライドし、収縮性タンパク質の空間構造の変化(らせんの形状の変化)によってそれらの短縮も可能であると示唆しています。
安静時、ATPには可塑化効果があります。ミオシンと組み合わせることにより、アクチンとの癒着の形成を防ぎます。 ATPは、筋肉の収縮中に分裂し、接着力の低下のプロセスと、Ca++イオンの供給である「カルシウムポンプ」の働きにエネルギーを提供します。 筋肉内のATPの分裂は、非常に高速で発生します。1分あたり1gの筋肉あたり最大10マイクロモルです。 筋肉内の総ATPリザーブは小さいので(最大パワーで0.5〜1秒の作業に十分です)、正常な筋肉活動を確保するために、ATPは分割されたのと同じ速度で回復する必要があります。
この教科書は、人体の筋肉活動の一般的な生化学と生化学の基礎を概説し、体の最も重要な物質の化学構造と代謝プロセスを説明し、筋肉活動を確保する上でのそれらの役割を明らかにします。 筋収縮のプロセスと筋肉のエネルギー生成のメカニズムの生化学的側面、運動の質の発達のパターン、疲労、回復、適応のプロセス、ならびに合理的な栄養とアスリートの機能状態の診断は次のとおりです。考慮。 体育とスポーツの高等教育機関の学生と教師のために、身体のリハビリテーションとレクリエーションの専門家。
書籍情報:
Volkov N.I.、Nesen E.N.、Osipenko A.A.、Korsun S.N. 筋活動の生化学。 2000.-503p。
パート1。 人体の生命活動の生化学的基盤
第1章生化学入門
1.生化学研究の主題と方法
2.生化学の発展とスポーツの生化学の形成の歴史
3.人体の化学構造
4.高分子の変換
テストの質問
第2章
1.代謝は生物の存在に必要な条件です
2.異化反応と同化反応-代謝の2つの側面
3.代謝の種類
4.細胞内の栄養素分解とエネルギー抽出の段階
5.細胞構造と代謝におけるそれらの役割
6.代謝の調節
テストの質問
第3章
1.エネルギー源
2.ATP-体内の普遍的なエネルギー源
3.生物学的酸化-体の細胞におけるエネルギー生産の主な方法
4.ミトコンドリア-細胞の「エネルギーステーション」
5.クエン酸回路は栄養素の好気性酸化の中心的な経路です
6.呼吸チェーン
7.酸化的リン酸化はATP合成の主なメカニズムです
8.ATP代謝の調節
テストの質問
第4章
1.水と体内でのその役割
2.水収支と筋肉活動中のその変化
3.ミネラルと体内でのそれらの役割
4.筋活動中のミネラルの代謝
テストの質問
第5章
1.物質輸送のメカニズム
2.体内環境の酸塩基状態
3.緩衝液システムと培地の一定のpHを維持する上でのそれらの役割
テストの質問
第6章
1.酵素の一般的な理解
2.酵素と補酵素の構造
3.複数の形態の酵素
4.酵素の性質
5.酵素の作用機序
6.酵素の作用に影響を与える要因
7.酵素の分類
テストの質問
第7章
1.ビタミンの一般的な理解
2.ビタミンの分類
3.脂溶性ビタミンの特性
4.水溶性ビタミンの特性評価
5.ビタミン様物質
テストの質問
第8章
1.ホルモンを理解する
2.ホルモンの性質
3.ホルモンの化学的性質
4.ホルモン生合成の調節
5.ホルモンの作用機序
6.ホルモンの生物学的役割
7.筋活動におけるホルモンの役割
テストの質問
第9章
1.炭水化物の化学組成と生物学的役割
2.炭水化物クラスの特性評価
3.人体の炭水化物の代謝
4.消化中の炭水化物の分解と血液への吸収
5.血糖値とその調節
6.炭水化物の細胞内代謝
7.筋活動中の炭水化物の代謝
テストの質問
第10章
1.脂質の化学組成と生物学的役割
2.脂質クラスの特性評価
3.体内の脂肪の代謝
4.消化中の脂肪の分解とそれらの吸収
5.細胞内脂肪代謝
6.脂質代謝の調節
7.脂質代謝の違反
8.筋活動中の脂肪の代謝
テストの質問
第11章
1.核酸の化学構造
2. DNAの構造、特性、生物学的役割
3. RNAの構造、特性および生物学的役割
4.核酸の交換
テストの質問
第12章
1.タンパク質の化学組成と生物学的役割
2.アミノ酸
3.タンパク質の構造的構成
4.タンパク質の性質
5.筋肉の働きに関与する個々のタンパク質の特性評価
6.遊離ペプチドと体内でのそれらの役割
7.体内のタンパク質代謝
8.アミノ酸の消化と吸収の間のタンパク質の分解
9.タンパク質生合成とその調節
10.間質性タンパク質の分解
11.アミノ酸の細胞内変換と尿素合成
12.筋活動中のタンパク質代謝
テストの質問
第13章代謝の統合と調節-適応プロセスの生化学的基礎
1.炭水化物、脂肪、タンパク質の相互変換
2.代謝の調節システムと身体の身体的ストレスへの適応におけるそれらの役割
3.中間代謝の統合における個々の組織の役割
テストの質問
パート2。 スポーツの生化学
第14章
1.筋肉と筋線維の種類
2.筋線維の構造組織
3.筋肉組織の化学組成
4.収縮および弛緩中の筋肉の構造的および生化学的変化
5.筋収縮の分子メカニズム
テストの質問
第15章
1.エネルギー生成メカニズムの一般的な特徴
2.ATP再合成のクレアチンホスホキナーゼメカニズム
3.ATP再合成の解糖メカニズム
4.ATP再合成のミオキナーゼメカニズム
5.ATP再合成の有酸素メカニズム
6.さまざまな物理的負荷時のエネルギーシステムの接続とトレーニング中のそれらの適応
テストの質問
第16章
1.筋活動中の生化学的プロセスの変化の一般的な方向
2.動作中の筋肉への酸素輸送と、筋肉活動中のその消費
3.筋肉の働きの間の個々の器官および組織の生化学的変化
4.筋肉の仕事中の生化学的変化の性質による身体運動の分類
テストの質問
第17章
1.最大および最大以下のパワーの短期運動中の疲労の生化学的要因
2.高および中程度のパワーの長期運動中の疲労の生化学的要因
テストの質問
第18章
1.筋肉の仕事の後の生化学的回復プロセスのダイナミクス
2.筋肉の働きの後のエネルギー貯蔵の回復の順序
3.筋肉の働きの後の休息期間中の崩壊生成物の除去
4.スポーツトレーニングの構築における回復プロセスの流れの特徴の使用
テストの質問
第19章
1.人の身体的パフォーマンスを制限する要因
2.アスリートの有酸素および無酸素パフォーマンスの指標
3.アスリートのパフォーマンスに対するトレーニングの効果
4.年齢とスポーツパフォーマンス
テストの質問
第20章
1.速度-強度品質の生化学的特性
2.アスリートのスピード-筋力トレーニングの方法の生化学的基盤
テストの質問
第21章
1.生化学的耐久性要因
2.持久力を促進するトレーニング方法
テストの質問
第22章
1.身体活動、適応およびトレーニング効果
2.生化学的適応の発達のパターンと訓練の原則
3.トレーニング中の身体の適応変化の特異性
4.トレーニング中の適応変化の可逆性
5.トレーニング中の適応変化のシーケンス
6.トレーニング中のトレーニング効果の相互作用
7.訓練の過程における適応の循環的発達
テストの質問
第23章
1.アスリートの合理的な栄養の原則
2.身体のエネルギー消費と実行された仕事への依存
3.アスリートの食事における栄養バランス
4.筋活動を確保する上での食品の個々の化学成分の役割
5.栄養補助食品と体重管理
テストの質問
第24章
1.生化学的管理のタスク、タイプ、および編成
2.研究対象と主な生化学的パラメーター
3.血液と尿の組成の主な生化学的指標、筋肉活動中のそれらの変化
4.筋活動中の体のエネルギー供給システムの発達の生化学的制御
5.アスリートの体のトレーニング、倦怠感、回復のレベルに対する生化学的制御
6.スポーツにおけるドーピングの管理
テストの質問
用語集
単位
文学
本についての詳細:フォーマット:pdf、ファイルサイズ:37.13Mb。
関連記事