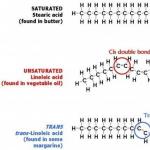
Sieviešu reproduktīvās sistēmas anatomija un fizioloģija īsumā. Sieviešu reproduktīvā sistēma: struktūra un fizioloģija. Sieviešu reproduktīvie orgāni
Sieviešu reproduktīvā funkcija tiek veikta galvenokārt olnīcu un dzemdes aktivitātes dēļ, jo olnīcas ir nobriedušas olu, un dzemdē olnīcu izdalīto hormonu ietekmē notiek izmaiņas, gatavojoties uztverei apaugļota olšūna. reproduktīvais periods ko raksturo sievietes ķermeņa spēja pavairot pēcnācējus; šī perioda ilgums ir no 17-18 līdz 45-50 gadiem. Pirms reproduktīvā jeb reproduktīvā perioda sievietes dzīvē ir šādi posmi: intrauterīns; jaundzimušais(līdz 1 gadam); bērnība(līdz 8-10 gadiem); pirmspubertātes Un puberitāte vecums (līdz 17-18 gadiem). Sākas reproduktīvais periods klimatiskais, kas atšķir premenopauze, menopauze Un pēcmenopauzes.
Menstruālais cikls - viena no sarežģītu bioloģisko procesu izpausmēm sievietes ķermenī. Menstruālo ciklu raksturo cikliskas izmaiņas visās reproduktīvās sistēmas daļās, kuru ārējā izpausme ir menstruācijas.
Menstruācijas -Šo asiņošana no dzimumorgānu trakta sievietēm, kas periodiski rodas no endometrija funkcionālā slāņa noraidīšanas divfāžu menstruālā cikla beigās. pirmās menstruācijas ( menarche) novēro 10-12 gadu vecumā, bet 1 - 1,5 gadu laikā pēc tam menstruācijas var būt neregulāras un pēc tam nostiprinās regulāras. menstruālais cikls.
Pirmā menstruāciju diena parasti tiek uzskatīta par menstruālā cikla pirmo dienu. Tāpēc cikla ilgums ir laiks starp nākamo divu menstruāciju pirmajām dienām. 60% sieviešu vidēji menstruālā cikla ilgums ir 28 dienas ar svārstībām no 21 līdz 35 dienām. Asins zuduma apjoms menstruāciju dienās 40-60 ml, vidēji 50 ml. Normālu menstruāciju ilgums no 2 līdz 7 dienām.
Olnīcas. Menstruālā cikla laikā olnīcas aug folikulu Un olu nobriešana, kas rezultātā kļūst gatavs apaugļošana. Tajā pašā laikā olnīcās tiek ražoti dzimumhormoni, kas nodrošina izmaiņas dzemdes gļotādā, kas spēj pieņemt apaugļotu olšūnu.
dzimumhormoni (estrogēni, progesterons, androgēni) ir steroīdi, piedalīties viņu izglītībā fanulārās šūnas folikuls, iekšējā un ārējā slāņa šūnas. dzimumhormoni olnīcu sintezētie ietekmē mērķa audus un orgānus. Tie ietver dzimumorgāni, Pirmkārt dzemde, piena dziedzeru, porains kauls, smadzenes, endotēlijs Un asinsvadu gludās muskulatūras šūnas, miokarda, āda un viņa pielikumi(matu folikuli un tauku dziedzeri) utt. Tiešs kontakts un specifiska hormonu saistīšanās ar mērķa šūnu ir tās mijiedarbības ar atbilstošajiem receptoriem rezultāts.
Bioloģisko efektu dod brīvās (nesaistītās) frakcijas estradiols Un testosterons(1 %). Lielākā daļa olnīcu hormonu (99%) ir saistītā stāvoklī. Transportēšanu veic īpaši proteīni - steroīdus saistošie globulīni un nespecifiskas transporta sistēmas - albumīns Un eritrocīti.
Zīmējums: Dominējošā folikula attīstības stadijas.
A - pirmatnējais folikuls; b - preantrālais folikuls; V - antrālais folikuls; G - preovulācijas folikuls: 1 - oocīts, 2 - granulozes šūnas (granulēta zona), 3 - theca šūnas, 4 - bazālā membrāna.
Estrogēnu hormoni dot ieguldījumu dzimumorgānu veidošanās, attīstība sekundārās seksuālās īpašības pubertātes laikā. Androgēni ietekmēt izskatu kaunuma un padušu apmatojums. Progesterons kontrolē menstruālā cikla sekrēcijas fāzi, sagatavo endometriju implantācijai. Dzimumhormoniem ir liela nozīme grūtniecības un dzemdību attīstībā.
Cikliskās izmaiņas olnīcās ietver trīs galvenos procesus:
1. Folikulu augšana un dominējošā folikulu veidošanās.
- Ovulācija.
- Dzeltenā ķermeņa veidošanās, attīstība un regresija.
Piedzimstot meitenei, olnīcās ir 2 miljoni folikulu, no kuriem 99% tiek pakļauti atrēzija visa mūža garumā. Atrēzijas process attiecas uz folikulu apgriezto attīstību vienā no tās attīstības posmiem. Tajā laikā menarche olnīcā ir aptuveni 200-400 tūkstoši folikulu, no kuriem 300-400 nobriest līdz ovulācijas stadijai.
Ir ierasts atšķirt šādus galvenos folikulu attīstības posmus: pirmatnējais folikuls, preantrālais folikuls, antrālais folikuls, preovulācijas folikuls.
Sākotnējais folikuls l sastāv no nenobriedušas olšūnas, kas atrodas folikulu un granulu (granulu) epitēlijā. Ārpus folikulu ieskauj savienojošs apvalks ( theca šūnas). Katra menstruālā cikla laikā sāk augt un veidoties 3 līdz 30 pirmatnējie folikuli pirmsantrāls, vai primārs, folikuli.
preantrālais folikuls. Ar izaugsmes sākumu pirmatnējais folikuls progresē uz preantrālo stadiju, un oocīts izplešas un to ieskauj membrāna, ko sauc spīdīgs apvalks (zona pellucida). Granulozes epitēlija šūnas proliferējas, un no apkārtējās stromas veidojas tekas slānis. Šo izaugsmi raksturo estrogēna ražošanas palielināšanās. Preantrālā folikula granulozes slāņa šūnas spēj sintezēt steroīdi trīs klases, savukārt estrogēni tiek sintezēti daudz vairāk nekā androgēni un progesterons.
Antral, vai sekundārais, folikuls . To raksturo turpmāka augšana: šūnu skaits granulozes slānī palielinās, ražojot folikulu šķidrums. Folikulārais šķidrums uzkrājas granulētā slāņa starpšūnu telpā un veido dobumus. Šajā folikuloģenēzes periodā (menstruālā cikla 8.-9. diena) tiek atzīmēta dzimumhormonu, estrogēnu un androgēnu sintēze.
Saskaņā ar mūsdienu teoriju par dzimumhormonu sintēzi androgēni tiek sintezēti teka šūnās - androstenedions Un testosterons. Pēc tam androgēni nonāk granulozes slāņa šūnās, kur aromatizējas par estrogēniem.
dominējošais folikuls . Parasti viens šāds folikuls veidojas no daudziem antrālajiem folikuliem (līdz cikla 8. dienai). Tas ir lielākais, satur visvairāk granulozes slāņa šūnu un FSH, LH receptorus. Dominējošajam folikulam ir bagātīgi vaskularizēts tekas slānis. Līdz ar dominējošā preovulācijas folikula augšanu un attīstību olnīcās paralēli notiek atlikušo (90%) augošo folikulu atrēzijas process.
Menstruālā cikla pirmajās dienās dominējošā folikula diametrs ir 2 mm, kas 14 dienu laikā līdz ovulācijas brīdim palielinās līdz vidēji 21 mm. Šajā laikā folikulu šķidruma tilpums palielinās 100 reizes. Tas strauji palielina estradiola un FSH saturu, un tiek noteikti arī augšanas faktori.
Ovulācija - preovulārā dominanta plīsums(terciārais) folikuls un olšūnas atbrīvošana no tā. Līdz ovulācijas brīdim olšūnā notiek process mejoze. Ovulāciju pavada asiņošana no salauztiem kapilāriem, kas ieskauj tekas šūnas. Tiek uzskatīts, ka ovulācija notiek 24-36 stundas pēc estradiola preovulācijas maksimuma veidošanās. Fermenta ietekmē notiek preovulācijas folikula sienas retināšana un plīsums kolagenāzes. Viņiem arī ir sava loma prostaglandīni F2a un Er, kas atrodas folikulu šķidrumā; granulozes šūnās ražoti proteolītiskie enzīmi; oksitocīns Un atslābina.
Pēc olšūnas izdalīšanas iegūtie kapilāri ātri ieaug folikula dobumā. Granulozes šūnas tiek pakļautas luteinizācija: tie palielina citoplazmas tilpumu un veido lipīdu ieslēgumi. LH, mijiedarbojoties ar granulozes šūnu proteīnu receptoriem, stimulē to luteinizācijas procesu. Šis process noved pie veidošanās dzeltenais ķermenis.
dzeltenais ķermenis - pārejošs endokrīnais dziedzeris, kas darbojas 14 dienas neatkarīgi no menstruālā cikla ilguma. Ja nav grūtniecības, dzeltenais ķermenis regresē.
Tādējādi galvenie sieviešu dzimuma steroīdie hormoni tiek sintezēti olnīcās - estradiols Un progesterons, un androgēni.
IN Menstruālā cikla I fāze, kas ilgst no pirmās menstruācijas dienas līdz ovulācijas brīdim, organisms atrodas estrogēnu ietekmē, un II (no ovulācijas līdz menstruāciju sākumam) tiek pievienots estrogēns. progesterons ko izdala dzeltenā ķermeņa šūnas. Pirmo menstruālā cikla fāzi sauc arī par folikulārs, vai folikulārs, cikla otrā fāze - luteāls.
Menstruālā cikla laikā perifērajās asinīs tiek atzīmēti divi estradiola satura maksimumi: pirmais ir izteikts preovulācijas cikls, bet otrs, mazāk izteikts, menstruālā cikla otrās fāzes vidū. Pēc ovulācijas cikla otrajā fāzē galvenais ir progesterons, kura maksimālais daudzums tiek sintezēts 4-7 dienā pēc ovulācijas.
Cikliskā hormonu sekrēcija olnīcā nosaka izmaiņas dzemdes gļotādā.
Cikliskas izmaiņas dzemdes gļotādā (endometrijā). Endometrijs sastāv no šādiem slāņiem:
Bāzes slānis , kas netiek noraidīts menstruāciju laikā. No tā šūnām menstruālā cikla laikā veidojas endometrija slānis.- Virsmas slānis, kas sastāv no kompaktām epitēlija šūnām, kas izklāj dzemdes dobumu.
- Vidēja līmeņa, vai porains, slānis.
Pēdējie divi slāņi veido funkcionālo slāni, kurā notiek lielas cikliskas izmaiņas menstruālā cikla laikā un izdalās menstruāciju laikā.
Menstruālā cikla I fāzē endometrijs ir plāns slānis, kas sastāv no dziedzeriem un stromas. Izšķir šādas galvenās endometrija izmaiņu fāzes cikla laikā:
1) proliferācijas fāze;
2) sekrēcijas fāze;
3) menstruācijas.
Proliferācijas fāze . Tā kā estradiola sekrēcija palielinās, augot olnīcu folikuliem, endometrijā notiek proliferatīvas izmaiņas. Notiek aktīva bazālā slāņa šūnu reprodukcija. Veidojas jauns virspusējs irdens slānis ar iegareniem cauruļveida dziedzeriem. Šis slānis ātri sabiezē 4-5 reizes. Cauruļveida dziedzeri, izklāta ar kolonnu epitēliju, iegareni.
Sekrēcijas fāze . Olnīcu cikla luteālajā fāzē progesterona ietekmē palielinās dziedzeru līkumainība, un to lūmenis pakāpeniski paplašinās. Stromas šūnas, palielinoties apjomam, tuvojas viena otrai. Palielinās dziedzeru sekrēcija. Dziedzeru lūmenā tiek konstatēts liels sekrēta daudzums. Atkarībā no sekrēcijas intensitātes dziedzeri vai nu paliek ļoti savīti, vai arī iegūst zāģa zoba formu. Ir pastiprināta stromas vaskularizācija. Ir agrīna, vidējā un vēlīnā sekrēcijas fāze.
Menstruācijas . Šis endometrija funkcionālā slāņa noraidīšana. Smalkie mehānismi, kas ir menstruāciju rašanās un procesa pamatā, nav zināmi. Ir noskaidrots, ka menstruāciju sākuma endokrīnais pamats ir izteikta progesterona un estradiola līmeņa pazemināšanās dzeltenā ķermeņa regresijas dēļ.
Ir šādi galvenie vietējie menstruāciju mehānismi:
1) spirālveida arteriolu tonusa izmaiņas;
2) izmaiņas hemostāzes mehānismos dzemdē;
3) izmaiņas endometrija šūnu lizosomu funkcijās;
4) endometrija reģenerācija.
Ir konstatēts, ka pirms menstruāciju sākuma notiek intensīva spirālveida arteriolu sašaurināšanās, kas izraisa išēmiju un deskvamācija endometrijs.
Saturs mainās menstruālā cikla laikā lizosomas endometrija šūnās. Lizosomas satur fermentus, no kuriem daži ir iesaistīti prostaglandīnu sintēzē. Reaģējot uz progesterona līmeņa pazemināšanos, šo enzīmu sekrēcija palielinās.
Endometrija reģenerācija novērota jau no paša menstruāciju sākuma. Līdz 24. menstruācijas stundas beigām tiek noraidītas 2/3 endometrija funkcionālā slāņa. Bāzes slānis satur stromas epitēlija šūnas, kas ir pamats endometrija reģenerācijai, kas parasti tiek pabeigta līdz cikla 5. dienai. Paralēli gali angioģenēze ar plosīto arteriolu, vēnu un kapilāru integritātes atjaunošanu.
Izmaiņas olnīcās un dzemdē notiek menstruālo funkciju regulējošo sistēmu divfāžu darbības ietekmē: smadzeņu garoza, hipotalāmu, hipofīze. Tādējādi tiek izdalītas 5 galvenās sieviešu reproduktīvās sistēmas saites: smadzeņu garoza, hipotalāmu, hipofīze, olnīcu, dzemde. Visu reproduktīvās sistēmas daļu savstarpējo savienojumu nodrošina gan dzimuma, gan gonadotropo hormonu receptoru klātbūtne tajās.
CNS loma reproduktīvās sistēmas darbības regulēšanā ir zināma jau sen. Tas tika pierādīts ovulācijas traucējumi ar dažādu akūtu un hronisku stresu, menstruālā cikla pārkāpums mainot klimatiskās un ģeogrāfiskās zonas, darba ritmu; plaši pazīstams menstruāciju pārtraukšana kara laikā. Garīgi nelīdzsvarotām sievietēm, kuras kaislīgi vēlas pēc bērna, var apstāties arī menstruācijas.
smadzeņu garozā un ekstrahipotalāmas smadzeņu struktūras(limbiskā sistēma, hipokamps, amigdala u.c.) ir noteikti specifiski estrogēnu, progesterona un androgēnu receptori. Šajās struktūrās notiek sintēze, izdalīšanās un metabolisms. neiropeptīdi, neirotransmiteri un to receptori, kas savukārt selektīvi ietekmē sintēzi un izdalīšanos hipotalāmu atbrīvojošais hormons.
Saistībā ar dzimumsteroīdu darbību viņarotransmiteri : norepinefrīns, dopamīns, gamma-aminosviestskābe, acetilholīns, serotonīns Un melatonīns. Norepinefrīns stimulē atbrīvošanos gonadotropīnu atbrīvojošais hormons (GTRG) no priekšējā hipotalāma neironiem. Dopamīns Un serotonīns samazināt frekvenci un samazināt ražošanas amplitūdu GTRG dažādās menstruālā cikla fāzēs.
Neiropeptīdi(endogēnie opioīdu peptīdi, neiropeptīds Y, kortikotropīnu atbrīvojošais faktors un galanīns) ietekmē arī reproduktīvās sistēmas darbību un līdz ar to arī hipotalāmu. Endogēni opioīdu peptīdi trīs veidi ( endorfīni, enkefalīni Un dinorfīni) spēj saistīties ar opiātu receptoriem smadzenēs. Endogēni opioīdu peptīdi ( Attēla pastiprinātājs) modulēt dzimumhormonu ietekmi uz saturu GTRG ar atgriezeniskās saites mehānismu bloķē gonadotropo hormonu sekrēciju no hipofīzes, īpaši LG, bloķējot GTRH sekrēciju hipotalāmā.
Mijiedarbība neirotransmiteri Un neiropeptīdi nodrošina regulārus ovulācijas ciklus sievietes organismā reproduktīvā vecumā, ietekmējot GTRH sintēzi un izdalīšanos hipotalāmā.
Hipotalāmā ir peptiderģiski neironi, kas izdala stimulējošus ( liberāļi) un bloķēšana ( statīni) neirohormoni - neirosekrēcija. Šīm šūnām piemīt gan neironu, gan endokrīno šūnu īpašības, un tās reaģē gan uz signāliem (hormoniem) no asinsrites, gan uz neirotransmiteriem un smadzeņu neiropeptīdiem. Neirohormoni tiek sintezēti neirona citoplazmas ribosomās un pēc tam tiek transportēti pa aksoniem uz galiem.
Gonadotropīnu atbrīvojošais hormons ( liberīns) ir neirohormons, kas regulē hipofīzes gonadotropo funkciju, kur tiek sintezēts FSH un LH. Atbrīvojošais hormons LH ( luliberīns) ir izolēts, sintezēts un detalizēti aprakstīts. Izolēt un sintezēt folikulus atbrīvojošo hormonu, vai folliberīns, līdz šim nav izdevies.
GnRH sekrēcijai ir pulsējošs raksturs: paaugstinātas hormona sekrēcijas maksimumi, kas ilgst vairākas minūtes, tiek aizstāti ar relatīvi zemas sekrēcijas aktivitātes 1-3 stundu intervāliem. GnRH sekrēcijas biežumu un amplitūdu regulē estrogēna līmenis.
Tiek saukts neirohormons, kas kontrolē prolaktīna sekrēciju ar adenohipofīzi prolaktīnu inhibējošais hormons(faktors), vai dopamīns.
Svarīga saikne reproduktīvajā sistēmā ir hipofīzes priekšējā daļa - adenohipofīze kurā izdalās gonadotropie hormoni folikulus stimulējošais hormons (FSH, folitropīns) luteinizējošais hormons (LG, lutropīns) Un prolaktīns (Prl), kas regulē olnīcu un piena dziedzeru darbību. Visi trīs hormoni ir olbaltumvielas ( polipeptīdi). Gonadotropo hormonu mērķa dziedzeris ir olnīca.
 Zīmējums: Reproduktīvās sistēmas funkcija (diagramma).
Zīmējums: Reproduktīvās sistēmas funkcija (diagramma).
RGLG - atbrīvo hormonus; labi - oksitocīns; Prl- prolaktīns; FSH - folikulus stimulējošais hormons; P - progesterons; E - estrogēni; A- androgēni; R - atslābina; UN - inhibīns; LG - luteinizējošais hormons.
Sintezē arī hipofīzes priekšējā daļā tirotropisks(TSH) Un adrenokortikotropisks(AKTH) hormoni, kā arī augšanas hormons.
FSH stimulē olnīcu folikulu augšanu un nobriešanu, veicina FSH un LH receptoru veidošanos uz olnīcu granulozes šūnu virsmas, palielina aromatāzes saturu nobriešanas folikulā un, stimulējot aromatizācijas procesus, veicina androgēnu pārvēršanos estrogēnos, stimulē inhibīna, aktivīna un insulīnam līdzīgā augšanas faktora-1 veidošanos, kam ir inhibējoša un stimulējoša loma folikulu augšanā.
L G stimulē:
androgēnu ražošana teka šūnās;
ovulācija ar FSH;
granulozes šūnu pārveidošana luteinizācijas laikā;
progesterona sintēze dzeltenajā ķermenī.
Prolaktīns stimulē piena dziedzeru augšanu un laktāciju, kontrolē progesterona sekrēciju dzeltenajā ķermenī, aktivizējot LH receptoru veidošanos tajos.
Prolaktīna sintēze, ko veic adenohipofīze, ir pakļauta dopamīna tonizējošas bloķēšanas kontrolei vai prolaktīna inhibējošais faktors. Prolaktīna sintēzes kavēšana grūtniecības un zīdīšanas laikā apstājas. Galvenais prolaktīna sintēzes stimulators ir tiroliberīns, kas sintezēts hipotalāmā.
Cikliskās izmaiņas hipotalāma-hipofīzes sistēmā un olnīcās ir savstarpēji saistītas un tiek modelētas kā atgriezeniskā saite.
Izšķir šādus atgriezeniskās saites veidus:
1) "gara cilpa"atsauksmes - starp olnīcu hormoniem un hipotalāma kodoliem; starp olnīcu un hipofīzes hormoniem;
2)"īsa cilpa"- starp hipofīzes priekšējo daļu un hipotalāmu;
3)"īpaši īsa cilpa"- starp GTRH un hipotalāma nervu šūnām.
Visu šo struktūru attiecības nosaka dzimumhormonu receptoru klātbūtne tajās.
Sievietei reproduktīvā vecumā ir gan negatīva, gan pozitīva atgriezeniskā saite starp olnīcām un hipotalāma-hipofīzes sistēmu. Negatīvas atsauksmes piemērs ir palielināta LH sekrēcija no hipofīzes priekšējās daļas reaģējot uz zemu estradiola līmeni cikla agrīnajā folikulu fāzē. Pozitīvu atsauksmju piemērs ir LH pieaugums reaģējot uz ovulācijas maksimumu estradiola saturs asinīs.
Par reproduktīvās sistēmas stāvokli var spriest, novērtējot funkcionālās diagnostikas testus: bazālā ķermeņa temperatūra, skolēna simptoms Un kariopiknotiskais indekss.
Bāzes temperatūra mēra taisnajā zarnā no rīta pirms izkāpšanas no gultas. Ovulācijas menstruālā cikla laikā bazālā temperatūra cikla luteālajā fāzē paaugstinās par 0,4-0,6 ° C un ilgst visu otro fāzi (sk. Att.). Menstruāciju dienā vai dienu pirms tām bazālā temperatūra pazeminās. Grūtniecības laikā bazālās temperatūras paaugstināšanās ir saistīta ar hipotalāma termoregulācijas centra ierosmi progesterona ietekmē.
 Zīmējums: Taisnās zarnas temperatūra divfāžu ciklā. M - menstruācijas; OV - ovulācija.
Zīmējums: Taisnās zarnas temperatūra divfāžu ciklā. M - menstruācijas; OV - ovulācija.
skolēna simptoms atspoguļo izmaiņas dzemdes kakla gļotās. Estrogēnu ietekmē dzemdes kaklā uzkrājas caurspīdīgas stiklveida gļotas, kas izraisa dzemdes kakla ārējās atveres paplašināšanos. Maksimālais gļotu daudzums tiek novērots cikla pirmsovulācijas dienās, ārējā atvere kļūst tumša, atgādina zīlīti. Cikla otrajā fāzē progesterona ietekmē samazinās gļotu daudzums vai arī tās pilnībā izzūd. Gļotām ir grumbuļaina struktūra. Ir 3 skolēna simptomu pakāpes: +, ++, +++.
Kariopiknotiskais indekss . Olnīcu hormonu ietekmē cikliskas izmaiņas notiek arī maksts gļotādā, īpaši tās augšējā trešdaļā. IN maksts uztriepes var saturēt sekojošo plakanu stratificēto epitēlija šūnu veidi : A) keratinizējošs, b) starpposma, V) bazāls, vai atrofisks. Pirmā tipa šūnas sāk dominēt, palielinoties estrogēnu sekrēcijai no olnīcām. Pamatojoties uz šūnu elementu kvantitatīvo attiecību noteikšanu, var spriest par organisma piesātinājuma pakāpi ar estrogēniem hormoniem vai to nepietiekamību. Maksimālais keratinizējošo šūnu skaits tiek konstatēts pirmsovulācijas dienās - 80-88%, agrīnā proliferācijas fāzē - 20-40%, vēlīnā sekrēcijas fāzē - 20-25%.
Vai informācija ir nepilnīga? Izmēģiniet meklēt no google .
Reproduktīvās sistēmas fizioloģija
1. Vispārīgi noteikumi, seksuālā diferenciācija
2. Vīriešu reproduktīvās sistēmas fizioloģija.
3. Sieviešu reproduktīvās sistēmas fizioloģija.
4. Olnīcu-menstruālais cikls.
Reproduktīvo funkciju regulēšana.
6. Grūtniecības fizioloģija.
7. Augļa fizioloģija.
8. Sievietes ķermeņa fizioloģija dzemdībās un pēcdzemdību periodā.
Vispārīgi noteikumi, seksuālā diferenciācija
Stāvs- organisma ģenētisko, morfoloģisko, fizioloģisko, psiholoģisko un sociāli personīgo īpašību kopums, kas nosaka tā specifisko līdzdalību vairošanās procesos.
Reproduktīvās funkcijas:
1) dzimumšūnu nobriešana;
2) seksuālā motivācija;
3) dzimumtieksme;
4) seksuālā uzvedība;
5) dzimumakts;
6) apaugļošanas process;
7) grūtniecība;
9) laktācija;
10) pēcnācēju barošana un audzināšana.
Orgāni un audi, kas veic šīs funkcijas, kā arī tos regulējošie mehānismi (nervu un humorālie) ir reproduktīvā sistēma. Viņas darbības gala rezultāts ir veselīgu pēcnācēju pavairošana.
Seksa pazīmes ir noteiktas embrionālais periods, tomēr dzimušais bērns ir nenobriedis. Savas dzīves laikā viņš iet cauri Vairāki seksuālās attīstības posmi:
1) bērnu stadija (līdz 8-10 gadiem);
2) pubertātes stadija (meitenes - 8-12 gadi, zēni - 10-14 gadi);
3) jaunības posms (meitenes - 13-16 gadi, zēni - 15-18 gadi);
4) pubertātes stadija (sievietēm - no 16-18 gadiem, vīriešiem - no 18-20 gadiem);
5) involūcijas stadija (sievietēm - pēc 45-55 gadiem, vīriešiem - pēc 60 gadiem).
Bērnība kam raksturīga nepilnīga dzimumdziedzeru un orgānu attīstība. Par to attīstību atbildīgo (gonadotropo), kā arī dzimumhormonu sekrēcija šajā periodā ir neliela.
IN puberitāte palielinās hipofīzes gonadotropo hormonu sekrēcija, sākas dzimumdziedzeru attīstība, paaugstinās dzimumhormonu līmenis asinīs. Uz šī fona notiek paātrināta primāro seksuālo īpašību attīstība un sākas sekundāro īpašību veidošanās.
Jauniešu posms ko raksturo straujš dzimumhormonu līmeņa paaugstināšanās asinīs. Apaugļošanās šajā vecumā jau ir iespējama, tomēr sievietes ķermeņa nepilnīga attīstība kļūst par grūtās grūtniecības un dzemdību gaitas cēloni. Kad tēvs ir jaunāks par 18 gadiem, palielinās risks piedzimt bērnam ar nepietiekamu ķermeņa masu. Turklāt ir grūtības ar adekvātu mazuļa audzināšanu.
Pubertātes laikā veidojas sekundārās seksuālās īpašības un fenotips. Androgēni ir tieši iesaistīti šajā procesā. Tie nosaka matu sadalījumu, ādas īpatnības, kaulu augšanu, muskuļu attīstību. Androgēni vīriešiem stimulē matu augšanu uz sejas, krūtīm un padusēs. Taču kombinācijā ar ģenētisko faktoru tiek nodrošināta pliku plankumu veidošanās temporālajos reģionos. Apmatojuma augšanu padusēs un kaunumā sievietēm nosaka arī androgēni. Androgēnu ražošanas palielināšanās sievietēm izraisa hirsutismu - pārmērīgu vīriešu matu augšanu.
Paaugstināta mērķa šūnu jutība pret androgēniem izraisa juvenīlu pūtīšu attīstību vīriešiem un sievietēm. Arī balsenes augšana un balss saišu sabiezēšana ir atkarīga no androgēniem, tāpēc einuhiem ir augsta balss toņa, tāpat kā zēniem pirms pubertātes. Androgēnu ietekmē aizveras kaulu epifīzes, kas neļauj indivīdam augt tālāk. Tāpēc priekšlaicīga pubertāte parasti tiek apvienota ar īsu augumu, savukārt cilvēki ar aizkavētu pubertāti un einuhi parasti ir gari.
pubertātes posms ko raksturo augsts dzimumhormonu līmenis, kā arī dzimumorgānu un dziedzeru galīgā attīstība, kas nodrošina organisma funkcionālo gatavību pavairot pilnvērtīgus pēcnācējus.
Involūcijas stadija ko raksturo pakāpeniska seksuālās funkcijas izzušana un dzimumhormonu līmeņa pazemināšanās asinīs. Spēja veikt dzimumaktu un dzimumtieksme ilgst daudz ilgāk nekā spēja apaugļot.
Involūcijas stadijas sākotnējais periods sauc par kulmināciju. Tas notiek gan vīriešiem, gan sievietēm, un to raksturo paaugstināta uzbudināmība, nogurums, garastāvokļa nestabilitāte, ko izraisa ierosmes un inhibīcijas procesi centrālajā nervu sistēmā. Šajā vecumā parādās vai saasinās dažādas slimības.
Tādējādi reproduktīvā sistēma nodrošina pilnvērtīgu pēcnācēju pavairošanu. Jaundzimušam bērnam tas ir nepilnīgs, bet pamazām cilvēks iziet cauri dažādiem dzimuma attīstības posmiem, kam raksturīgs atšķirīgs reproduktīvās sistēmas darbības līmenis.
Seksuālā diferenciācija
Primārās dzimumšūnas - gonocīti tiek izolēti embrija šūnās ļoti agrīnās attīstības stadijās (6. nedēļā). Tie tiek pārvietoti uz nākamo dzimumdziedzeru zonu, vispirms ar asins plūsmu caur embrija asinsvadiem un pēc tam pārvietojas neatkarīgi. Šajā posmā vīriešu un sieviešu gonocīti ir gandrīz vienādi, atšķirības parādās tikai pēc iekļūšanas dzimumdziedzeros.
Organisma dzimums, tāpat kā jebkura iezīme, attīstās, no vienas puses, genotipa un, no otras puses, vides faktoru ietekmē. Dažādiem organismiem genotipa un vides faktoru ietekme uz dzimuma noteikšanu ir atšķirīga, t.i. dažiem organismiem (cilvēkiem, vairumam zīdītāju) noteicošais ir genotips, citos (zivīs, atsevišķi tārpi) - vides faktori. Tātad tārpā Bonellia viridis mātīte ir salīdzinoši liela, tēviņš mazs. Viņš pastāvīgi dzīvo sievietes dzimumorgānos. Tārpa kāpurs ir divdzimumu, tēviņa vai mātītes attīstība no šāda kāpura ir atkarīga no gadījuma. Ja kūniņa, kādu laiku peldot ūdenī, satiekas ar mātīti, kas ir brīva no tēviņa un piefiksējas pie tās, tā pārvērtīsies par tēviņu, bet, ja nē, par mātīti.
Dažreiz vides faktori būtiski ietekmē dzimuma noteikšanu arī zīdītājiem. Tātad liellopiem, vienlaikus attīstoties diviem pretējā dzimuma dvīņiem, buļļi piedzimst normāli, un teles bieži ir starpdzimumu. Tas ir saistīts ar agrāku vīriešu dzimuma hormonu izdalīšanos un to ietekmi uz otrā dvīņa dzimumu.
Dzimuma maiņu var novērot Atlantijas siļķēs. Reņģes dzīvo nelielos saimēs, no kuriem katrā ir viens tēviņš un vairākas mātītes. Ja tēviņš nomirst, tad pēc kāda laika lielākā mātīte pārvēršas par tēviņu.
Cilvēkiem ir aprakstīti vīriešu fenotipa izpausmes gadījumi ar dzimuma hromosomu saturu XX un sievietes (Morisa sindroms) - ar XY genotipu. Ar Morisa sindromu embrioģenēzes laikā tiek novietoti sēklinieki, kas sāk ražot vīriešu dzimuma hormonus. Taču šādi embriji neveido receptorproteīnu (recesīvu gēna mutāciju), kas nodrošina jaunattīstības orgānu šūnu jutību pret vīrišķo dzimumhormonu. Sakarā ar to apstājas attīstība atbilstoši vīrieša tipam un parādās sievietes fenotips.
Nonākuši dzimumdziedzeru rudimentos, abu dzimumu gonocīti intensīvi vairojas ar parastu mitotisku dalījumu. Embrijam veidojas nediferencētu dzimumdziedzeru rudimentu pāris – dzimumorgānu krokas. Viņi vienmēr ir klāt, neatkarīgi no nedzimušā bērna dzimuma. Seksuālo diferenciāciju nosaka dzimuma hromosomu sastāvs. Tie satur informāciju par proteīna sintēzi, kas stimulē dzimumorgānu pamatu attīstību. Ja augļa genotips satur Y-hromosomu, sākas aktīva testosterona sintēze. Tas mijiedarbojas ar īpašiem receptoriem mērķa šūnās un stimulē to dzimumorgānu kroku daļu attīstību, kas izraisa vīriešu reproduktīvo sistēmu. Ja uz vīrieša genotipa fona tiek traucēta šo receptoru jutība vai izkropļota testosterona ražošana, reproduktīvā sistēma attīstās atbilstoši sievietes tipam.
Sieviešu dzimumdziedzeru diferenciācija šajā periodā ir vāji izteikta. Testosterona trūkums ļauj rudimentiem attīstīties sieviešu veidā. Dzimumdziedzerī notiek primāro dzimumšūnu mitoze un veidojas folikulu sākumi.
Tādējādi dzimumdziedzeri sākotnēji tiek likti neatkarīgi no embrija dzimuma. Y-hromosoma, kas ir atbildīga par testosterona sintēzi, kļūst par izšķirošu faktoru attīstībā. Testosterona klātbūtnē rudimenti attīstās atbilstoši vīrieša tipam, ja nav - pēc mātītes.
Reproduktīvie orgāni attīstās no divām struktūrām: Millera un Volffa kanāliem.
Ieslēgts agrīnās stadijas tie atrodas visos embrijos neatkarīgi no dzimuma. Androgēnu iedarbībā vīriešu dzimuma auglim no Volffa kanāla attīstās epididīms, vas deferens un sēklas pūslīši. Millera kanālu inhibējošais faktors veicina Millera kanāla atrofiju.
Sieviešu auglim deģenerējas Volffa kanāls, un no Millera kanāla attīstās olšūns, dzemde, dzemdes kakls un maksts augšdaļa.
Sievietes auglim urīnizvadkanāla krokas neaug kopā, bet veido mazās kaunuma lūpas. Lielās kaunuma lūpas veidojas no pāra izciļņiem. Seksuālais tuberkuls tiek pārveidots par klitoru. Šo struktūru, kā arī iekšējo dzimumorgānu attīstība notiek neatkarīgi no olnīcām.
Vīrieša auglim, lai nediferencēti rudimenti pārvērstos ārējos dzimumorgānos, asinīs ir nepieciešams pietiekams daudzums androgēnu. Viņu darbības rezultātā urīnizvadkanāla krokas aug kopā, veidojot sēklinieku maisiņu. Dzimumorgānu tuberkulozes izmērs palielinās, pārvēršoties par dzimumlocekli.
Embrioģenēzes procesā dzimumorgānu rudimenti sākotnēji atrodas blakus nierēm un pēc tam migrē uz leju. Olnīcas paliek iegurņa dobumā, un sēklinieki nolaižas sēklinieku maisiņā. Viņu klātbūtne ir ārkārtīgi svarīga, jo normālai testosterona ražošanai un pilnvērtīgai spermatoģenēzei ir nepieciešama temperatūra, kas ir nedaudz zemāka par ķermeņa temperatūru. Ja sēklinieki nenolaižas sēkliniekos, vīrietis paliek sterils.
Tādējādi ārējo un iekšējo dzimumorgānu attīstība galvenokārt ir atkarīga no androgēnu klātbūtnes vai neesamības, kas nosaka dzimuma attīstības veidu.
VĪRIEŠU REĢENERĀLĀ SISTĒMA
Vīriešu reproduktīvie orgāni
Vīriešu reproduktīvie orgāni tiek iedalīti ārējos (sēklinieku maisiņi, dzimumloceklis) un iekšējos (sēklinieki ar piedēkļiem, vas deferens, prostatas dziedzeris, bulbourethral dziedzeri, sēklas pūslīši un vas deferens). Divi sēklinieki tiek iznesti ārpus kaunuma kaula un karājas sēklinieku maisiņā. Sēklinieks sastāv no piramīdveida lobulām, no kurām katra satur vītņotas un taisnas sēklinieku kanāliņus. Sēklinieks ir savienots ar epididīmu, kas to ieskauj, ar līdz 6 m garu saritinātu cauruli un vas deferens, kas iet uz prostatas dziedzeri. Pirms iekļūšanas prostatas dziedzerī vas deferens savienojas ar sēklas pūslīšu izvadkanālu. Vas deferens beigu posma un sēklas pūslīšu ekskrēcijas kanāla saplūšanas rezultātā veidojas ejakulācijas kanāls. Ejakulācijas kanāls perforē prostatas dziedzeri un atveras urīnizvadkanālā (urethra). Ārpus prostatas bulbourethral (Cooper) dziedzeri atveras urīnizvadkanālā. Dzimumlocekļa ķermenī ir kavernozi un poraini ķermeņi. Dzimumlocekļa porainajā ķermenī atrodas urīnizvadkanāls, kas beidzas ar dzimumlocekļa galviņu.
sēklinieks - saplacinātas-ovālas formas pāra orgāns, 4 cm garš un 2,5 cm diametrā Sēklinieks ar piedēkli atrodas sēkliniekos - maisiņā, kas atrodas ārpus vēdera dobuma tieši aiz dzimumlocekļa. Iekšējo slāni, kas izklāj sēklinieku maisiņa dobumu (sēklinieku maisiņa viscerālo slāni), sauc par maksts membrānu (tunica vaginalis). T. vaginalis ir vēderplēves slānis, kas pārvietojas jaunattīstības sēklinieku maisiņā. Šajā gadījumā vēderplēves izvirzīšanas rezultātā caur vēdera priekšējo sienu veidojas iegarena cauruļveida kabata, ko veido vēderplēve - maksts process (processus vaginalis), pa kuru migrē sēklinieks. Pēc sēklinieka pārvietošanas sēklinieku maisiņā processus vaginalis aizaug.
Lielāko daļu sēklinieku veido vītņoti kanāliņi, kas satur spermatogēno epitēliju. Izliektie kanāliņi, tuvojoties sēklinieku videnei, pārvēršas taisnos kanāliņos, kas savukārt pāriet tīkla kanāliņos, kas atrodas tieši sēklinieku videnē. Taisni un izliekti kanāliņi kalpo spermatozoīdu izvadīšanai, kas veidojas tikai vītņoto sēklu kanāliņu spermatogēnajā epitēlijā.
PIELIKUMS sēklinieku(epididymis) ir komata forma, atrodas blakus sēklinieka posterolaterālajai virsmai un sastāv no ārkārtīgi un haotiski izlocītas caurules, kuras garums ir līdz 6 m, ko sauc par epididimijas kanālu (ductus epididymidis). Sākot no epididimijas galvas, kas atrodas uz sēklinieka augšējā pola, d. epididymidis veido piedēkļa ķermeni un asti. Piedēkļa astes apakšējā daļā d. epididymidis pāriet tiešās vas deferens - ductus (vas) deferens.
SEMINĀLS AUKTS. Visi sēklinieku un epididimijas asinsvadi un limfātiskie asinsvadi no vēdera dobuma caur cirkšņa kanālu nonāk sēklinieku maisiņā, kopā ar ductus deferens un to pavadošajām nervu šķiedrām, kā arī membrānām, kas stiepjas no vēdera priekšējās daļas. siena, tā sauktais spermatozoīds (funiculus spermaticus).
SĒĶKOKU KANĀLS- epididimijas kanāliņa turpinājums - 45 cm caurule, kas atkāpjas no epididimijas apakšējā gala un paceļas gar sēklinieka aizmuguri. Vas deferens kā daļa no spermas vada nonāk vēdera dobumā, kur tas atrodas gar iegurņa iekšējo sienu. Tuvojoties sēklas pūslīšiem, kanāls izplešas (ampulla) un savienojas ar sēklas pūslīšu kanālu, veidojot īsu (2,5 cm) ejakulācijas kanālu (ductus ejaculatorius), kas ieplūst urīnizvadkanāla prostatas daļā.
SĒKLAS BURBUĻI- divi ļoti izliekti kanāliņi, kuru garums ir līdz 15 cm, kas atrodas urīnpūšļa pamatnē taisnās zarnas priekšpusē.
PROSTATA DZIEZERES(prostata) - dziedzeru-muskuļu orgāns ar izmēru 2-4 cm, kas ieskauj vīrieša urīnizvadkanāla sākotnējo posmu, t.i. atrodas vietā, kur tā iziet no urīnpūšļa. Prostatas parenhīma sastāv no 30–50 sazarotiem cauruļveida alveolāriem dziedzeriem. Dziedzeru kanāli atveras urīnizvadkanāla prostatas daļā.
SEKSUĀLS BIEDRS. Galvenā dzimumlocekļa masa ir erekcijas audi, kas sakārtoti 3 struktūru veidā, kas atrodas visā orgāna garumā. Pārī savienoti, cilindriski kavernozi ķermeņi (corpora cavernosa) atrodas dzimumlocekļa muguras pusē, bet ventrālajā - porains ķermenis (corpus spongiosum). Dzimumlocekļa virsotne (galva) ir porainā ķermeņa paplašinātā distālā daļa. Erekcijas audu pārplūšana ar asinīm izraisa ievērojamu dzimumlocekļa izmēra palielināšanos un tā iztaisnošanu - erekciju. Dzimumlocekļa galva ir pārklāta ar plānu ādu, tās apļveida kroku, kas aptver galvu, sauc par priekšādiņu. Dzimumlocekļa inervāciju, kas ir ļoti svarīga erekcijai, veic pudendālais nervs (S 2–4) un iegurņa pinumi.
SPERMATOĢĒZE
spermatoģenēze tiek veikta īpašās struktūrās, ko sauc par vītņotām sēklu kanāliņām, kurām ir ļoti izliekta gaita un kas atrodas sēklinieku daivu iekšpusē. Epitēlijs, kas tos pārklāj, sastāv no jaunattīstības spermatozoīdiem un atbalsta šūnām. Šo epitēliju sauc par spermatogēnu. Sēklinieku šķērsgriezumos redzami spermatocīti dažādos nobriešanas posmos. Starp spermatogēnajām šūnām ir Sertoli šūnas, kuru funkcijas ir: trofisks(nodrošinot jaunattīstības gametas ar barības vielām), fagocitoze liekā spermatīda citoplazma un deģenerējošas dzimumšūnas, aromatizēšana androgēni (testosterona pārvēršana par estrogēniem, kas nepieciešama endokrīno Leidiga šūnu funkciju lokālai regulēšanai), sekrēcijušķidrums un androgēnu saistošais proteīns (nepieciešams spermatozoīdu transportēšanai sēklu kanāliņos) un endokrīnais (inhibīnu sintēze). Svarīga Sertoli šūnu funkcija ir hematotestikulārās barjeras izveide.
Intersticijā starp vītņotajām sēklu kanāliņām atrodas Leidiga šūnas, kuru funkcijas ir androgēnu (testosterona, dihidrotestosterona, dehidroepiandrosterona, androstenediona un dažu citu) ražošana.
Testosterons, tāpat kā citi androgēni, tas ir būtisks seksuālai diferenciācijai, pubertātei, sekundāro seksuālo īpašību uzturēšanai un spermatoģenēzei (skatīt zemāk). Testosterons - anaboliskais hormons. Šajā kapacitātē dažādos orgānos (aknās, skeleta muskuļos, kaulos) testosterons stimulē olbaltumvielu sintēzi. Jo īpaši testosterona ietekmē palielinās muskuļu masa, blīvums un kaulu masa. Eritropoetīna sintēzes stimulēšanas rezultātā palielinās Hb un hematokrīta (Ht) saturs, un aknu lipāzes sintēzes palielināšanās asinīs izraisa augsta blīvuma lipoproteīnu līmeņa pazemināšanos asinīs un zema blīvuma lipoproteīnu satura palielināšanās. Citiem vārdiem sakot, testosteronam ir izteikta aterogēna iedarbība, t.i. veicina aterosklerozes (tostarp koronāro asinsvadu) attīstību.
Vīriešiem spermatoģenēzes process ilgst 65-70 dienas. Tas notiek visā sēklu kanāliņos. Jauns cikls sākas ar vienādiem laika intervāliem, tāpēc katrā kanāliņā var redzēt šūnas dažādās attīstības stadijās. Tādā veidā tiek uzturēta ilgstoša nepārtraukta spermatozoīdu ražošana. Katru dienu veidojas apmēram 2 x 10 8. Spermatogonija vīrieša ķermenī turpina dalīties no pubertātes sākuma līdz vecumam.
spermatozoīdi - mazas šūnas, to diametrs ir 1-2 mikroni. To forma ir labi pielāgota kustībai un mijiedarbībai ar olu. Mejozes rezultātā no katra spermatogonija veidojas četri identiski spermatozoīdi. Spermatozoīda galvā ir kodols, kas satur haploīdu hromosomu skaitu. To klāj akrosoma, kas ir īpaša ar membrānu saistīta struktūra, kas satur hidrolītiskos enzīmus. Fermenti atvieglo spermas iekļūšanu olšūnā tieši pirms apaugļošanas. Funkcionāli to dažreiz uzskata par paplašinātu lizosomu.
Šķidrums, kas ejakulēts dzimumakta laikā (ejakulāts) - sperma, tas satur vīriešu reproduktīvās sistēmas palīgdziedzeru (sēklu pūslīšu, prostatas un bulbouretrālo dziedzeru) spermatozoīdus un sekrēcijas šķidrumu. Sēklu šķidrumā spermatozoīdi veido 5% no tilpuma, 95% - palīgdziedzeru noslēpumus.
Ejakulāta daudzums katras kopulācijas laikā ir 3,5 (2-6) ml, katrā mililitrā ir aptuveni 120 miljoni spermatozoīdu. Lai nodrošinātu auglību (auglību), katrā spermas mililitrā ir jābūt vismaz 20 miljoniem spermatozoīdu (tostarp 60% no normālas morfoloģijas un vairāk nekā 50% no mobilajiem). Pēc ejakulācijas maksimālais spermatozoīdu dzīves ilgums sievietes dzimumorgānu traktā nepārsniedz 48 stundas. Tajā pašā laikā temperatūrā, kas zemāka par -100 ° C, spermatozoīdi gadiem ilgi paliek auglīgi.
sēklas pūslīši izdala viskozu, dzeltenīgu noslēpumu, kas ejakulācijas laikā nonāk ejakulācijas kanālā. Sēklu pūslīšu noslēpums atšķaida sēklu, satur fruktozi, askorbīnskābes un citronskābes sāļus, Pg - t.i. vielas, kas nodrošina spermu ar enerģijas rezervi, palielina to izdzīvošanu un funkcionālo aktivitāti.
Prostata. Dziedzera noslēpums piedalās sēklu sašķidrināšanā un atvieglo tās izkļūšanu caur urīnizvadkanālu ejakulācijas laikā. Dziedzera noslēpums satur bikarbonātu, lipīdus, proteolītiskos enzīmus (fibrinolizīnu), skābo fosfatāzi. Nedaudz sārmainā sekrēta reakcija (pH 7,5) neitralizē citu sēklu šķidruma sastāvdaļu skābumu un tādējādi palielina spermatozoīdu kustīgumu un auglību (apaugļošanas spēju). Prostata veic arī endokrīnās funkcijas, sintezējot bioloģiski aktīvās vielas, kas nomāc testosterona sekrēciju.
bulbourethral dziedzeri Kūpers. Seksuālās uzbudinājuma laikā izdalītais viskozs gļotādas sekrēts kalpo urīnizvadkanāla eļļošanai pirms ejakulācijas.
Vīrieša organismā notiek dažādi procesi (gan tieši saistīti ar reproduktīvo funkciju, gan nosakot vīriešu somatiskos, psiholoģiskos un uzvedības fenotipus) regulē androgēnu (steroīdie vīriešu dzimuma hormoni), inhibīnus, hipotalāma luliberīnu, hipofīzes gonadotropos hormonus (LH un FSH), kā arī estradiolu. un dažas citas bioloģiski aktīvas vielas.
GnRH sintezēts hipotalāma neirosekrēcijas šūnās. Sasniedzot hipofīzes priekšējās daļas hipotalāma-hipofīzes asinsrites sistēmu, GnRH aktivizē endokrīnās šūnas, kas sintezē FSH un LH.
Gonadotropie hormoni(folikulu stimulējošā - FSH un luteinizējošā - LH) tiek ražoti adenohipofīzē. To sekrēciju kontrolē gan GnRH ( aktivizējas), un sēklinieku hormoni ( apspiest). Gonadotropo hormonu mērķi - sēklinieki. Sertoli šūnām ir FSH receptori, savukārt Leidiga šūnām ir LH receptori.
FSH. Sertoli šūnas ir FSH mērķis izliektajos sēklu kanāliņos. FSH receptoru stimulēšana noved pie intracelulāro androgēnu receptoru sintēzes un androgēnu saistoša proteīna veidošanās, kas saista Leidiga šūnu ražoto testosteronu un transportē to uz spermatogēnām šūnām. Turklāt Sertoli šūnas izdala inhibīnus, kas kopā ar testosteronu kavē FSH veidošanos.
LG stimulē Leidiga šūnas ražot testosteronu. Papildus LH receptoriem Leidiga šūnām ir receptori prolaktīns Un inhibīni. Šie hormoni pastiprina LH stimulējošo iedarbību uz testosterona veidošanos, bet bez LH testosterona sintēze nenotiek.
Testosterons. Galvenais spermatoģenēzes aktivators.
Estrogēni. Sertoli šūnās, aromatizējot, Leidiga šūnās sintezētais testosterons tiek pārveidots par estrogēnu. Lai gan šis ieguldījums estrogēna līmenī asinīs ir neliels, Sertoli šūnām ir būtiska ietekme uz testosterona sintēzi. Estrogēni saistās ar receptoriem Leidiga šūnās un kavē testosterona sintēzi. Turklāt estrogēni samazina gonadotropo šūnu jutību pret GnRH.
Inhibīni. Reaģējot uz FSH stimulāciju, Sertoli šūnas izdala inhibīnus, kas bloķē FSH un GnRH sintēzi un sekrēciju. Inhibīnu struktūra ir homologa Miller inhibējošajam faktoram, ko izdala augļa Sertoli šūnas.
SIEVIEŠU REĢENERĀLĀ SISTĒMA
Sieviešu reproduktīvo sistēmu veido pārī savienotas olnīcas un olvados, dzemde, maksts, ārējie dzimumorgāni un piena dziedzeri. Orgānu struktūra un funkcijas atšķiras. Tātad, olnīcu funkcijas - dīgstošs( ovoģenēze, ovulācija) un endokrīnās sistēmas(estrogēnu, progesterona, relaksīnu un inhibīnu sintēze un sekrēcija), olvadi - transports(ovulētas olšūnas ievadīšana dzemdes dobumā, apaugļošana), dzemde - grūtniecība, dzemdes kakla kanāls un maksts - dzimšanas kanāls piena dziedzeri ir būtiski, lai bērna barošana.
olnīcasir sieviešu dzimumdziedzeri. Tie atrodas iegurņa dobumā pie sānu sienām. Sievietēm nobriedušā vecumā vidējie olnīcu izmēri ir šādi: garums - 3-4 cm, platums - 2-2,5, biezums - 1-1,5 cm, svars - 6-8 g Olnīcā, dzemdē un olvados izšķir galus.. Caurules gals ir pacelts uz augšu un vērsts pret dzemdes (olvadas) piltuvi. Olnīca ir kustīgi savienota ar saitēm ar dzemdes un iegurņa sieniņu.
Dzemdeir bumbierveida forma, kas vērsta pret maksts augšējās daļas šauro galu. Dzemdē izšķir dibenu, ķermeni, kaklu un dobumu. Apakšdaļa ir dzemdes augšējā daļa virs olvados. Ķermenim ir trīsstūra forma, tā turpinājums, kas veido apakšējo daļu, ir dzemdes kakls. Dzemdējas sievietes dzemdes dobumam frontālajā daļā ir trīsstūra forma. Šī trijstūra augšējos stūros ir atveres, kas atveras olvados, apakšējā stūrī ir šaurums, kas ved uz dzemdes kakla kanāla dobumu. Dzemdes kakls ir konisks vai cilindrisks. Tās apakšējā galā kanāls atveras makstī.
Maksts- muskuļu-elastīgā caurule, kas atrodas mazajā iegurnī, ar tās augšējo galu nosedz dzemdes kaklu, apakšējā beidzas maksts vestibilā. Jaunavām vestibila dibenu un tā apakšējo galu ierobežo jaunavības plēve. Virzoties no iegurņa dobuma uz vestibilu, maksts iet caur uroģenitālo diafragmu. Maksts ir iesaistīta kopulācijas un apaugļošanās procesos, dzemdībās tā ir daļa no dzemdību kanāla. Nobriedušas sievietes maksts garums svārstās no 7 līdz 9 cm, platums - 2-3 cm, aizmugurējā siena ir par 1,5-2 cm garāka nekā priekšējā. Maksts var mainīt savu formu, diametru un dziļumu, saraujoties iegurņa pamatnes muskuļiem, dzemdei un saišu aparāta muskuļu elementiem.
Funkcionāli maksts ir sadalīta divās daļās: augšējā un apakšējā augšējā daļa ir paplašināta, tā spēj aktīvi sarauties, apakšējā ir sašaurināta un masīvāka.
Seksuālās uzbudinājuma periodā maksts vēnās asinis piepildās, pagarinās tās augšējās daļas, palielinās transudācija maksts lūmenā. Pēc dzimumakta maksts gļotāda spēj absorbēt spermas plazmu un prostaglandīnus, ko ražo sēklas pūslīši. Dzemdību laikā maksts ir stipri izstiepta, bet nedēļu pēc tām sieniņu elastības dēļ maksts saraujas, lai gan tās lūmenis paliek plašāks nekā pirms dzemdībām.
Uz leju no uroģenitālās diafragmas, kas aizver izeju no mazā iegurņa, atrodas ārējie sieviešu dzimumorgāni. Tajos ietilpst sieviešu dzimumorgānu zona (vulva). Sieviešu dzimumorgānu apvidū ietilpst kaunums, lielās un mazās kaunuma lūpas, klitors, maksts vestibils, tā dziedzeri, vestibila spuldze. Dzimumorgānu sadalījums ārējos un iekšējos ir skaidrojams ne tikai ar to topogrāfijas īpatnībām, bet arī ar embrionālās attīstības un funkciju specifiku. Sieviešu dzimumorgānu attīstība daļēji notiek rumpja apakšējās daļas ādas dēļ.
Pubis ir zemākā vēdera sienas daļa. Tam ir trīsstūra forma, kuras pamatne ir vērsta uz leju. Pubis pāriet lielajās kaunuma lūpās. Lielās kaunuma lūpas ir sapārotas parasagitāli izvietotas ādas izciļņi, kuru biezumā ir iestrādāti taukaudi ar venozo pinumu un tajā ietvertiem elastīgo šķiedru kūļiem. Mazās kaunuma lūpas atrodas mediāli no lielajām un tām paralēli. To biezumā ir arī saistaudi un salīdzinoši liels venozais pinums. Kopā ar lielajām kaunuma lūpām tie ierobežo dzimumorgānu spraugu no sāniem. Dzimumorgānu spraugas priekšējā stūrī starp mazajām kaunuma lūpām atrodas klitors, kura biezumā atrodas kavernozs ķermenis. Nedaudz aiz klitora, starp to un maksts ieeju, atrodas urīnizvadkanāla ārējā atvere, kas atveras maksts vestibilā. Vestibila dibenu veido himēns. Himēna pamatā ir saistaudi ar elastīgām, kolagēna un muskuļu šķiedrām, kas veido tās turgoru. Lielo kaunuma lūpu pamatnē un biezumā ir novietotas divas nepāra kavernoza veidojuma daivas - vestibila sīpoli.
Klitors satur lielu skaitu mehānoreceptoru. Seksuālās uzbudinājuma laikā klitors uzbriest. Tas ir saistīts ar palielinātu arteriālo asins plūsmu un samazinātu venozo aizplūšanu. Paralēli tam uzbriest vestibila spuldze, kas ir venozs pinums, kas atgādina kavernozu ķermeni. Šajā brīdī no vestibila dziedzeriem izdalās ar mucīnu bagāts noslēpums, kas mitrina maksts ieeju.
Grūtniecības fizioloģija.
MĀLĪBA
Olas apaugļošana parasti notiek dzemdes (olvados) vadā - sapārotā cauruļveida orgānā, kas veic olšūnas un spermas transportēšanas funkcijas, radot labvēlīgus apstākļus apaugļošanai, olšūnas attīstībai grūtniecības sākumā un embrija attīstīšanai pirmajās grūtniecības dienās. attīstība dzemdē. Olvads vienā galā atveras dzemdē, otrs - vēderplēves dobumā pie olnīcām. Vēdera atvere, kuras diametrs ir 2-3 mm, parasti ir aizvērta. Tās atklāšana ir saistīta ar ovulācijas procesu. Ovulācijas laikā olvadu vēdera gals var būt ciešā saskarē ar olnīcu. Olvadā ir izolēta piltuve, ampula un šaurums. Piltuve atveras vēderplēves dobumā, tās bārkstiņas satver olšūnu ovulācijas laikā un vēl vairāk veicina virzību ampulā. Ampula ir tieši tā vieta, kur notiek apaugļošanās. Tam ir vāji izteikts muskuļu slānis un augsti attīstīts epitēlijs. Straume atrodas caurules un dzemdes savienojuma vietā un ir dobs lūmenis, kas ir mehānisks šķērslis šūnu kustībai.
Olvados dzimumšūnas tiek transportētas pretējos virzienos. Spermatozoīdi pārvietojas no dzemdes uz ampulu, un zigotas, kas rodas pēc apaugļošanas, pārvietojas dzemdes dobumā. Gludo muskuļu kontrakciju koordinācijai un skropstu kustības pakāpei ir nepieciešama smalka koordinācija, kas tiek panākta ar īpašām hormonālām un neirālām ietekmēm.
apaugļošana To sauc par spermas saplūšanu ar olšūnu, kā rezultātā veidojas zigota, kas var augt, attīstīties un radīt jaunu organismu. Apaugļošanas laikā vīrišķo un sieviešu dzimumšūnu kodolmateriāls apvienojas, kas noved pie tēva un mātes gēnu apvienošanās, hromosomu diploīdā komplekta atjaunošanas.
Cilvēkiem ejakulāts tiek ievietots makstī. Tās tilpums ir 2-5 ml un satur no 30 līdz 100 miljoniem spermatozoīdu uz 1 ml. Tomēr tikai daži miljoni no tiem iekļūst dzemdes kakla kanālā tā dobumā, un tikai aptuveni 100 spermatozoīdu sasniedz olvadu augšējo daļu. Spermatozoīdi, kas paliek makstī, nevar tur ilgstoši pastāvēt skābās vides (pH 5,7) dēļ, lai gan zināmu aizsardzību šajā gadījumā nodrošina ejakulāta sārmainās īpašības. Dzemdes dobumā spermas izdzīvošanas apstākļi arī nav tik labvēlīgi, bet cita iemesla dēļ. Šeit liela nozīme ir leikocītu augstajai fagocītiskajai aktivitātei. Turklāt viens no šķēršļiem spermatozoīdu virzībā uz olnīcām ir mehāniskās kustības grūtības dzemdes olvadu reģionā. Tam visam kopumā ir sava pozitīvā puse, kas neļauj novājinātām vai neparastām dzimumšūnām iekļūt olvados. Izdzīvojušie spermatozoīdi var sasniegt olvadu ampulu 10-20 minūšu laikā pēc dzimumakta. Tik strauju progresu nevar nodrošināt tikai ar spermas kustīgumu. Veicināšanu veicina vairāki faktori, tostarp maksts muskuļu kontrakcijas, miometrija kontrakcijas, ciliāru kustības, peristaltikas kontrakcijas un šķidruma plūsma olvados. Dažos gadījumos spermas šūnas šķērso visu olvadu un apaugļo olšūnu tūlīt pēc ovulācijas, pirms tā nonāk olšūnas piltuvē. Šādos gadījumos embrijs var piestiprināties pie olnīcas vai vēdera sienām, izraisot attīstību. ārpusdzemdes grūtniecība.
Periods, kurā spermatozoīdi mātītes dzimumorgānos saglabā spēju apaugļot, ir salīdzinoši īss: pelē - 6 stundas, jūrascūciņā - 22 stundas, trusis - līdz 36 stundām. Sievietēm dzimumorgānu traktā spermatozoīdi saglabā spēju apaugļot 2-4 dienas. Dzīvniekiem ir izņēmumi. Tātad dažiem sikspārņiem pārošanās notiek rudenī, un olu ovulācija un to apaugļošana notiek tikai pavasarī. Tādējādi viņu spermatozoīdi saglabā spēju apaugļot vairākus mēnešus.
Apaugļošana ietver šādus procesus: olšūnas atpazīšana ar spermatozoīdu; spermas iekļūšanas olšūnā regulēšana, polispermijas novēršana; otrā meiotiskā dalījuma beigas; vīriešu un sieviešu priekškodolu veidošanās, šūnu dalīšanās sākums.
Atpazīšanas procesu raksturo vairāki mehānismi, un, pirmkārt, ir zināms, ka olšūnas caurspīdīgās membrānas glikoproteīni darbojas kā spermatozoīdu receptori. Šie receptori ir ļoti specializēti un specifiski sugai. Tas pilnībā izslēdz jebkādu dzimumšūnu saplūšanu starp sugām.
Spermatozoīda iekļūšana olšūnā sākas ar lielu kontaktu skaitu starp plazmas membrānu un spermatozoīda akrosomālo membrānu. Mijiedarbības rezultātā parādās pūslīši ar proteolītiskajiem enzīmiem. Šie fermenti tikai izšķīdina folikulu šūnu matricu un caurspīdīgo membrānu. Spermatozoīds iekļūst kanālā, kas veidojas fermentatīvās darbības dēļ caurspīdīgajā apvalkā, izmantojot astes dzinējspēku.
Polispermijas novēršana tiek panākta arī ar vairākiem mehānismiem, no kuriem galvenais ir tas, ka tūlīt pēc pirmā spermatozoīda iekļūšanas (iekļūšanas) notiek gandrīz tūlītēja olšūnas membrānas depolarizācija, kas pārvēršas par pastāvīgu blokādi (process ir pētīts). detalizēti jūras eži). Pilnīga blokāde rodas, aktivizējot kortikālās granulas, kas ir lizosomu organellas, kas satur proteolītiskos enzīmus. Granulu saturs tiek ielejams pericelulārajā telpā un iekļūst caurspīdīgajā membrānā. Rezultātā spermas receptori tiek inaktivēti, savukārt pati caurspīdīgā membrāna kļūst blīva un nepieejama turpmākai vīrišķo dzimumšūnu iejaukšanās gadījumā.
Spermatozoīda un olšūnas saplūšana izraisa ienākošo kalcija jonu strāvu un kalcija izdalīšanos no intracelulārajām depo, tādējādi aktivizējot apaugļoto olšūnu (zigotu). Izmantojot virkni starpposma mehānismu, zigota nonāk pirmajā mitotiskajā nodaļā. Divu šūnu veidošanās stadija ilgst no 24 līdz 36 stundām.
Zigota, kas izveidojusies pēc apaugļošanas, pamazām virzās uz dzemdi un nonāk tajā pēc dažām dienām. 2-3 dienu laikā tas atrodas dzemdes dobumā suspendētā stāvoklī. Ēdienu nodrošina tur esošais šķidrums. Zigotas piestiprināšana (implantācija) pie dzemdes sienas notiek tikai 6-7 dienā pēc ovulācijas. Šajā periodā dzemdes sienas endometrijs estrogēnu un progesterona iedarbības rezultātā tiek sagatavots implantācijas procesam.
Ovulāciju, apaugļošanu un implantāciju var ietekmēt vairāki līdzekļi un kontracepcijas metodes(aizsardzība no ieņemšanas). Tas arī šeit ir īsi jāatzīmē, jo pēdējam procesam ir ievērojama praktiska nozīme.
Priekšmeta "Iegurnis no dzemdniecības viedokļa. Sieviešu reproduktīvās sistēmas fizioloģija" satura rādītājs:1. Iegurnis no dzemdniecības viedokļa. Iegurņa dobums.
2. Mazā iegurņa platās daļas plaknes izmēri. Mazā iegurņa šaurās daļas plaknes izmēri.
3. Iegurņa stieples ass. Iegurņa leņķis.
5. Olnīcas. Cikliskas izmaiņas olnīcās. Primordiāls, preantrāls, antrāls, dominējošais folikuls.
6. Ovulācija. dzeltens ķermenis. Olnīcās sintezētie sieviešu hormoni (estradiols, progesterons, androgēni).
7. Cikliskas izmaiņas dzemdes gļotādā (endometrijā). proliferācijas fāze. sekrēcijas fāze. Menstruācijas.
8. Centrālās nervu sistēmas loma menstruāciju regulēšanā. Neirohormoni (luteinizējošais hormons (LH), folikulus stimulējošais hormons (FSH).
9. Atgriezeniskās saites veidi. Atgriezeniskās saites sistēmas loma menstruālo funkciju regulēšanā.
10. Bāzes temperatūra. skolēna simptoms. Kariopiknotiskais indekss.
Sieviešu reproduktīvā funkcija tiek veikta galvenokārt olnīcu un dzemdes aktivitātes dēļ, jo olšūna nogatavojas olnīcās, un dzemdē olnīcu izdalīto hormonu ietekmē notiek izmaiņas, gatavojoties apaugļotas augļa olšūnas uztverei, reproduktīvo periodu raksturo sievietes ķermeņa spēja atražot pēcnācējus; šī perioda ilgums ir no 17-18 līdz 45-50 gadiem.
Pirms reproduktīvā jeb reproduktīvā perioda sievietes dzīvē ir šādi posmi: intrauterīns; jaundzimušie (līdz 1 gadam); bērnība (līdz 8-10 gadiem); pirmspubertātes un pubertātes vecums (līdz 17-18 gadiem). Reproduktīvais periods pāriet menopauzē, kurā ir premenopauze, menopauze un pēcmenopauze.
Menstruālais cikls- viena no sarežģītu bioloģisko procesu izpausmēm sievietes ķermenī. Menstruālais cikls ko raksturo cikliskas izmaiņas visās reproduktīvās sistēmas daļās, kuru ārējā izpausme ir menstruācijas.
Mēs iesakām noskatīties apmācības video: Rīsi. Cikliskas izmaiņas reproduktīvās sistēmas orgānos menstruālā cikla laikā.I - olnīcu funkcijas gonadotropā regulēšana;
PDH - hipofīzes priekšējā daļa;
II - estradiola receptoru saturs endometrijā - RE (1,2,3; nepārtraukta līnija) un progesterons - RP (2,4,6; punktēta līnija);
III - cikliskas izmaiņas endometrijā;
IV - maksts epitēlija citoloģija;
V - bazālā temperatūra;
VI - dzemdes kakla gļotu sasprindzinājums.
Menstruācijas- tie ir asiņaini izdalījumi no sievietes dzimumorgānu trakta, kas periodiski rodas no endometrija funkcionālā slāņa noraidīšanas divfāžu menstruālā cikla beigās. Pirmās menstruācijas (menarhe) tiek novērotas 10-12 gadu vecumā, bet 1-1,5 gadu laikā pēc tām menstruācijas var būt neregulāras, un tad tiek noteikts regulārs menstruālais cikls.
Pirmā menstruāciju diena nosacīti pieņemts kā menstruālā cikla pirmā diena. Tāpēc cikla ilgums ir laiks starp nākamo divu menstruāciju pirmajām dienām. 60% sieviešu vidējais menstruālā cikla ilgums ir 28 dienas, ar svārstībām no 21 līdz 35 dienām. Asins zuduma apjoms menstruāciju dienās ir 40-60 ml, vidēji 50 ml. Normālu menstruāciju ilgums ir no 2 līdz 7 dienām.
Dzimumorgāni veic reproduktīvo funkciju, tie nosaka cilvēka seksuālās īpašības. Gan vīriešiem, gan sievietēm dzimumorgānus iedala iekšējos un ārējos.
Vīriešu reproduktīvie orgāni
Pie vīriešu dzimumorgāniem pieder sēklinieki ar to piedēkļiem, sēklinieku dobumi un ejakulācijas vadi, sēklas pūslīši, prostatas un bulbouretrālie dziedzeri, sēklinieki un dzimumloceklis (4.5. att.).
Iekšējie vīriešu reproduktīvie orgāni.sēklinieki, vai sēklinieki- tvaika vīrišķais dziedzeris, kura funkcija ir vīriešu dzimumšūnu - spermatozoīdu veidošanās un vīrišķo dzimumhormonu izdalīšanās asinīs.
Sēklinieki ir ovāli, 4,5 x 3 cm lieli, sver 20–30 g; tie atrodas sēkliniekos, un kreisais sēklinieks ir zemāks par labo. Sēkliniekus vienu no otra atdala sēklinieku maisiņš, un tos ieskauj membrānas. Sēklinieks ir apturēts uz spermas auklas, kas ietver asinsvadus, muskuļus un fasciju, asins un limfas asinsvadus un nervus.
Blakus sēklinieka aizmugurējai malai atrodas piedēklis, kurā izšķir galvu, ķermeni un asti.
Rīsi. 4.5. Vīriešu reproduktīvie orgāni
Vēderplēve aptver sēklinieku no visām pusēm un veido slēgtu serozu dobumu. Ārpusē sēklinieks ir pārklāts ar baltu šķiedru membrānu, ko sauc balts apvalks, zem kura ir sēklinieku parenhīma. No albudžijas aizmugurējās malas iekšējās virsmas saistaudu izaugums nonāk sēklinieku parenhīmā - sēklinieku videnes, no kuriem ir plānas sēklinieku saistaudu starpsienas, kas sadala dziedzeri daudzos (no 250 līdz 300) piramīdveida lobulas, virzīts ar galotnēm uz sēklinieku videnes, un ar pamatni - uz albuginea. Katras daivas biezumā ir divas vai trīs vītņoti sēklu kanāliņi 60–90 mm garš, to ieskauj irdeni saistaudi un daudzi asinsvadi. Iekšpusē esošie sēklu kanāliņi ir izklāti ar stratificētu spermatogēno epitēliju, šeit veidojas vīriešu dzimumšūnas, spermatozoīdi. Pēdējās ir daļa no sēklas, kuras šķidrā daļa veidojas no sēklas pūslīšu un prostatas noslēpumiem. Sēklu kanāliņi saplūst, veidojot tiešie sēklu kanāliņi, kas ieplūst retikulā. No sēklinieku tīkla rodas 12-15 eferenti kanāliņi, kas iziet cauri albuginei un ieplūst epididimijas kanālā.
vas deferens- pāra orgāns, kura garums ir aptuveni 50 cm, diametrs ir 3 mm un lūmena diametrs ir aptuveni 0,5 mm. Pamatojoties uz kanāla topogrāfiskajām iezīmēm, tajā izšķir četras daļas: sēklinieku, kas atbilst sēklinieka garumam; aukla - spermas vadā; cirkšņa - cirkšņa kanālā un iegurņa - no dziļā cirkšņa gredzena līdz prostatas dziedzerim.
Pēc sēklu kanāla iziešanas vas deferens savienojas ar sēklas pūslīšu izvadkanālu ejakulācijas kanālā. Pēdējais caurums atveras urīnizvadkanāla prostatas daļā.
sēklas pūslīši- sapārots sekrēcijas orgāns 10–12 cm garš un 0,6–0,7 cm biezs Veziklas atrodas iegurņa dobumā urīnpūšļa sānos un aiz apakšas. Katrā sēklas pūslī ir pamatne (platais gals), korpuss (vidējā daļa) un apakšējais (šaurais) gals, kas nonāk izvadkanālā. Sēklas pūslīšu siena sastāv no gļotādas, muskuļu un nejaušām membrānām; tajā ir daudz līkumotu kameru, kas satur olbaltumvielu šķidrumu, kas ir daļa no spermas.
Prostata- nesapārots muskuļu un dziedzeru orgāns, kas sver 20–25 g, izdala noslēpumu, kas ir daļa no spermas. Tas atrodas zem urīnpūšļa mazā iegurņa apakšā (sk. 4.4. att.). Forma atgādina kastaņu, nedaudz saspiests anteroposterior virzienā.
Prostatas dziedzerim ir labās un kreisās daivas, šaurums; ārpusē tas ir pārklāts ar kapsulu, no kuras starpsienas nonāk orgāna iekšpusē. Tas sastāv no dziedzeru un gludu muskuļu audiem. Dziedzeru audi veido dziedzeru parenhīmu, un tos attēlo īpaši kompleksi alveolāru-cauruļveida lobulu veidā. Orgānu dziedzeru ejas nonāk prostatas ekskrēcijas kanālos, kas punktos atveras vīriešu urīnizvadkanāla lūmenā. Muskuļu audi aizpilda prostatas priekšējo daļu un, savienojoties ar urīnpūšļa dibena muskuļu saišķiem, veido urīnizvadkanāla iekšējo (piespiedu) sfinkteru.
Bulbourethral dziedzeris (Kūpera dziedzeris) - pārī savienots orgāns, kas atrodas aiz vīrieša urīnizvadkanāla membrānas daļas starpenes dziļā šķērseniskā muskuļa biezumā. Dziedzerim ir alveolāri cauruļveida struktūra, blīva tekstūra, ovāla forma, diametrs 0,3–0,8 cm.Urīnvadā atveras bulbouretrālo dziedzeru kanāli. Dziedzeris ražo viskozu šķidrumu, kas aizsargā urīnizvadkanāla sienas gļotādu no urīna izraisīta kairinājuma.
Ārējie vīriešu dzimumorgāni ko attēlo dzimumloceklis un sēklinieku maisiņš.
Dzimumloceklis- orgāns, kas kalpo urīna izvadīšanai un spermas izvadīšanai (4.6. att.). Tas sastāv no priekšējās brīvās daļas - ķermeņa, kas beidzas ar galvu, un muguras daļas, kas piestiprināta pie kaunuma kauliem. Dzimumlocekļa galvā izšķir platāko daļu - galvas vainagu un šauro - galvas kaklu.

Rīsi. 4.6. Dzimumlocekļa struktūra.
Dzimumlocekļa ķermeni klāj plāna, viegli kustīga āda. Tās apakšējā virsmā ir šuve. Ķermeņa priekšējā daļā veidojas ādas kroka - dzimumlocekļa priekšāda, kas nosedz glans, un pēc tam nonāk dzimumlocekļa galvas ādā. Orgāna apakšējā virsmā priekšāda ir savienota ar galvu ar priekšādas frenuluma palīdzību. Dzimumlocekļa augšdaļā atveras urīnizvadkanāla ārējā atvere, kas izskatās kā vertikāla sprauga.
Dzimumlocekļa ķermenis sastāv no diviem kavernoziem ķermeņiem un viena nepāra - poraina. porains ķermenis aizmugure beidzas ar spuldzi, bet priekšā - ar dzimumlocekļa galvu. Iekšpusē porains ķermenis iet caur urīnizvadkanālu, kas izplešas galvā un veido navikulāru fossa. Kavernozi ķermeņi ir cilindriska forma, to aizmugurējie gali novirzās uz sāniem dzimumlocekļa kāju veidā un ir piestiprināti pie kaunuma kaulu apakšējiem zariem.
Kavernozi un poraini ķermeņi sastāv no specifiskiem sūkļveida audiem un spēj savākt asinis savos daudzajos dobumos (dobumos) un kļūt diezgan blīvi; līdz ar asiņu aizplūšanu tie samazinās.
Sēklinieku maisiņš- muskuļu un ādas maisiņš, kurā atrodas sēklinieki un piedēkļi, kā arī spermatozoīdu auklu apakšējās daļas. Sēklinieku maisiņā ir septiņi slāņi (čaumalas): āda; gaļīgs apvalks; ārējās sēklas fascijas; muskuļa fascija, kas paceļ sēklinieku; muskulis, kas paceļ sēklinieku; iekšējā sēklas fascija un sēklinieku cirkšņa membrāna, kurā izšķir divas loksnes (parietālo un iekšējo). Sēklinieku maisiņa sienas čaumalas atbilst vēdera priekšējās sienas slāņiem, jo tās veidojas, sēkliniekam nolaižoties no vēdera dobuma sēkliniekos. Sēklinieku maisiņa dobums ir sadalīts divās daļās ar starpsienu; katra puse ir konteiners vienam sēkliniekam.
spermatoģenēze ir vīriešu dzimuma šūnu veidošanās process. Tas ir pirmais un galvenais rādītājs pubertātes sākumam jauniem vīriešiem un ilgst gandrīz visu mūžu. Spermatoģenēze sastāv no trim posmiem un notiek vīriešu dzimumdziedzeru sēklinieku kanāliņos - sēkliniekos (sēkliniekos) (4.7. att.).

Rīsi. 4.7. Spermatoģenēzes stadijas un spermas struktūra
Pirmais posms ir daudzās spermas šūnu mitozes; otrais ir mejoze; trešais ir spermioģenēze. Sākotnēji veidojas spermatogonijas, kas atrodas uz spermatozoīdu vadu ārējās sienas. Tad tie secīgi pārvēršas par pirmās kārtas spermatocītiem. Pēdējie, daloties meiotiski, dod divas identiskas šūnas - otrās kārtas spermatocītus. Otrās dalīšanas laikā otrās kārtas spermatocīti rada četras nenobriedušas dzimumšūnas - gametas. Tos sauc par spermatīdiem. Iegūtie četri spermatīdi pakāpeniski pārvēršas par aktīviem, kustīgiem spermatozoīdiem.
Sieviešu reproduktīvie orgāni
Sieviešu reproduktīvo sistēmu veido ārējie un iekšējie dzimumorgāni, un to raksturo primārās un sekundārās sievietes īpašības. Tajā ietilpst olnīcas un to piedēkļi, dzemde un olvadi, maksts, klitors un sievietes dzimumorgānu zona (4.8. att.).
Ārējie sieviešu dzimumorgāni. Tie atrodas priekšējā starpenē uroģenitālā trīsstūrī un ietver sieviešu dzimumorgānu zonu un klitoru.
UZ sieviešu dzimumorgānu zona ietver kaunumu, lielās un mazās kaunuma lūpas, maksts vestibilu, vestibila lielākos un mazākos dziedzerus un vestibila spuldzi.
Pubis augšpusē to no vēdera atdala kaunuma rievas, bet no gurniem - gurnu rievas. Tas ir pārklāts ar matiem, kas stiepjas līdz lielajām kaunuma lūpām. Zemādas tauku slānis ir labi attīstīts kaunuma zonā.
Lielas kaunuma lūpas tās ir 7–8 cm garas un 2–3 cm platas noapaļotas pāra krokas, kas ierobežo dzimumorgānu spraugu no sāniem. Lielas kaunuma lūpas savā starpā ir savienotas ar priekšējo un aizmugurējo komisāru. Āda, kas aptver lielas kaunuma lūpas, satur daudz tauku un sviedru dziedzeru.

Rīsi. 4.8 . Sieviešu reproduktīvie orgāni
Starp lielajām kaunuma lūpām ir vēl viens pāris ādas kroku - mazās kaunuma lūpas. To priekšējie gali aptver klitoru, veido klitora priekšādiņu un frenulumu, un aizmugurējie gali, savienojoties kopā, veido šķērsvirziena kroku - kaunuma lūpu frenulumu. Atstarpi starp mazajām kaunuma lūpām sauc maksts vestibils. Tas satur urīnizvadkanāla ārējo atveri un maksts atveri.
Klitors ir vīriešu dzimumlocekļa kavernozo ķermeņu homologs un sastāv no pārī savienotiem kavernozs ķermeņiem. Tas atšķir ķermeni, galvu un kājas, kas piestiprinātas kaunuma kaulu apakšējiem zariem. Priekšpusē klitora ķermenis sašaurinās un beidzas ar galvu. Klitoram ir blīva šķiedraina albuginea, un to klāj āda, kas bagāta ar maņu nervu galiem.
Starpene - mīksto audu komplekss (āda, muskuļi, fascijas), kas aizver ieeju no iegurņa dobuma. Tas aizņem apgabalu, ko priekšā ierobežo kaunuma simfīzes apakšējā mala, aiz kaunuma kaula gals un sānos – kaunuma un sēžas kaula apakšējie zari un sēžamvietas bumbuļi. Līnija, kas savieno sēžas bumbuļus, sadala starpenumu divos trīsstūros: priekšējo augšējo daļu sauc. uroģenitālā, un muguras lejasdaļa anālā zona. Uroģenitālajā reģionā atrodas uroģenitālā diafragma, bet tūpļa - iegurņa diafragma.
Uroģenitālā diafragma un iegurņa diafragma ir muskuļu-fasciāla plāksne, ko veido divi muskuļu slāņi (virspusēja un dziļa) un fascija.
Virspusējie muskuļi Uroģenitālās diafragmas ietver virspusējos šķērseniskos starpenes, ischiocavernosus un bulbospongius muskuļus. UZ dziļie muskuļi Uroģenitālā diafragma ietver dziļo šķērsenisko starpenes muskuļu un urīnizvadkanāla sfinkteru.
IN iegurņa diafragma nokļūst virspusējais muskuļu slānis, ko attēlo nepāra muskulis - tūpļa ārējais sfinkteris. Saraujoties, tas saspiež (aizver) tūpļa atveri. Iegurņa diafragmas dziļajos muskuļos ietilpst divi muskuļi, kas veido iegurņa dobuma dibena aizmugurējo daļu: muskuļu, kas paceļ tūpļa un coccygeal.
Sievietes kājstarpe atšķiras no vīrieša. Uroģenitālā diafragma sievietēm ir plaša, caur to iet urīnizvadkanāls un maksts; muskuļi ir nedaudz vājāki nekā vīriešiem, un fascijas, gluži pretēji, ir stiprākas. Urīnizvadkanāla muskuļu saišķi pārklāj arī maksts sienu. Starpenes cīpslu centrs atrodas starp maksts un anālo atveri, sastāv no cīpslām un elastīgajām šķiedrām.
Iekšējie sieviešu reproduktīvie orgāni.Olnīca- tvaika sieviešu dzimumdziedzeris, kas atrodas iegurņa zonā (4.9. att.). Olnīcu masa ir 5-8 g; garums 2,5–5,5 cm, platums 1,5–3,0 cm, biezums līdz 2 cm. Olnīca ir olveida, nedaudz saspiesta anteroposterior virzienā. Ar savu un piekares saišu palīdzību tas tiek fiksēts abās dzemdes pusēs. Fiksācijā piedalās arī vēderplēve, kas veido olnīcas apzarni (dublēšanos) un piestiprina to pie dzemdes platās saites. Olnīcā izšķir divas brīvas virsmas: mediāla, kas vērsta mazā iegurņa dobumā, un sānu, blakus mazā iegurņa sienai. Olnīcu virsmas aiz muguras nonāk izliektā brīvā (aizmugurējā) malā, priekšā - mezenteriskajā malā, pie kuras ir piestiprināta olnīcas apzarnis.
Mezenteriskās malas reģionā ir depresija - olnīcu vārti caur kuriem ieplūst un iziet asinsvadi un nervi. Olnīcā izšķir augšējo olvadu galu, kas ir pagriezts pret olvadu, un apakšējo dzemdes galu, kas savienots ar dzemdi ar savu olnīcas saiti. Šī saite atrodas starp diviem plašās dzemdes saites slāņiem. Olnīcas olvadu lielākā olnīcu fimbrija ir piestiprināta pie olnīcas olvadu gala.
Olnīcas ir iekļautas mobilo orgānu grupā, to topogrāfija ir atkarīga no dzemdes stāvokļa, tās lieluma.
Olnīcu virsma ir pārklāta ar vienu dīgļu epitēlija slāni, zem kura atrodas blīvs saistaudu albuginea. Iekšējā viela (parenhīma) ir sadalīta ārējā un iekšējā slāņos. Olnīcu ārējo slāni sauc kortikālā viela. Tas satur lielu skaitu folikulu, kas satur olas. Starp tiem ir vezikulāri olnīcu (nobrieduši) folikuli (grafijas pūslīši) un nobriest primārie olnīcu folikuli. Nobriedis folikuls var būt 0,5–1,0 cm liels; pārklāts ar saistaudu membrānu, kas sastāv no ārējā un iekšējā slāņa.

Rīsi. 4.9. Iekšējie sieviešu reproduktīvie orgāni.
Dzemde- dobs, nepāra orgāns, kurā notiek embrija attīstība un augļa nēsāšana. Tas atšķir apakšējo daļu - augšējo daļu, ķermeni - vidējo daļu un kaklu - apakšējo sašaurināto daļu. Tiek saukta sašaurināta dzemdes ķermeņa pāreja uz dzemdes kaklu dzemdes sašaurinājums. Dzemdes kakla apakšējo daļu, kas nonāk maksts dobumā, sauc dzemdes kakla maksts daļa, un augšējais, kas atrodas virs maksts, - supravaginālā daļa. Dzemdes atvēršanu ierobežo priekšējās un aizmugurējās lūpas. Aizmugurējā lūpa ir plānāka nekā priekšējā. Dzemdei ir priekšējā un aizmugurējā virsma.
Dzemdes izmērs un svars atšķiras. Dzemdes garums pieaugušai sievietei ir vidēji 7–8 cm, bet biezums – 2–3 cm. Tas atrodas iegurņa dobumā starp taisno zarnu un urīnpūsli.
Dzemde tiek fiksēta ar kreiso un labo plato saišu palīdzību, kas sastāv no diviem vēderplēves slāņiem (priekšējā un aizmugurējā). Dzemdes platās saites laukumu, kas atrodas blakus olnīcai, sauc olnīcu apzarnis. Dzemdi tur arī apaļās saites un dzemdes kardinālās saites.
Dzemdei lielā mērā ir mobilitāte, kas ir atkarīga no blakus esošo orgānu stāvokļa.
Olvads (olvads).– pāra cauruļveida orgāns 10–12 cm garš, 2–4 mm diametrā; veicina olšūnas pāreju no olnīcas uz dzemdes dobumu. Olvadi atrodas abās dzemdes dibena pusēs, ar šauru galu atveras dzemdes dobumā, bet ar paplašinātu - vēderplēves dobumā. Tādējādi caur olvadām vēderplēves dobums ir savienots ar dzemdes dobumu.
Olvadā izšķir piltuvi, ampulu, šauru un dzemdes daļu. Piltuve ir caurules ventrālā atvere, kas beidzas ar garām šaurām bārkstīm. Piltuvei seko olvadu ampula, tālāk - tā šaurā daļa - isthmus. Pēdējais iet uz dzemdes daļa, kas caur olvadu atveras dzemdes dobumā.
Maksts- nesapārots dobs orgāns 8–10 cm garas caurules veidā, sienas biezums 3 mm. Ar augšējo galu tas aptver dzemdes kaklu, un ar apakšējo galu caur iegurņa uroģenitālo diafragmu tas atveras vestibilā ar maksts atveri. Šo caurumu jaunavai aizver himēns, kas ir pusmēness vai perforēta plāksne, kas tiek saplēsta pirmā dzimumakta laikā, un pēc tam tās atloki atrofējas. Maksts priekšā ir urīnpūslis un urīnizvadkanāls, aiz - taisnā zarna, ar kuru tā saplūst ar vaļīgiem un blīviem saistaudiem.
Maksts priekšējās un aizmugurējās sienas ir izolētas, kas ir savienotas viena ar otru. Nosedzot dzemdes kakla maksts daļu, tie ap to veido kupolveida ieplaku - maksts velve.
Ooģenēze - sieviešu dzimumšūnu attīstības process olnīcā. Primārās sieviešu dzimuma šūnas (oogonia) sāk attīstīties pirmajos intrauterīnās attīstības mēnešos. Pēc tam oogonija pārvēršas par oocīti. Līdz dzimšanas brīdim meiteņu olnīcās ir aptuveni 2 miljoni olšūnu, kas pārvēršas par pirmās kārtas olšūnām. Tomēr starp tiem ir intensīvs atrēzijas process, kas ievērojami samazina to skaitu. Pirms pubertātes sākuma paliek apmēram 500 000 oocītu, kas spēj tālāk dalīties. Pēc tam oocīti attīstās pirmatnējos folikulos un pēc tam pirmatnējos folikulos. Sekundārie folikuli parādās tikai pēc pubertātes sasniegšanas.
Sekundārais folikuls turpina augt un pārvēršas par nobriedušu (grafijas pūslīšu). Tad folikuls plīst un olšūna nonāk vēderplēves dobumā (4.10. att.). Šo procesu sauc ovulācija.

Rīsi. 4.10. Olu nobriešanas un ovulācijas process:
1 - olvads, 2 - olnīca, 3 - dzemde, 4 - bālgans ķermenis (deģenerēts dzeltenais ķermenis), 5 - primārie folikuli, 6 - barojošie asinsvadi, 7 - olšūna, 8 - nobriest folikuls, 9 - nobriedusi olšūna (ovulācija) , 10 - nobriedis dzeltenais ķermenis, 11 - attīstās dzeltenais ķermenis
Sievietes menstruālo (seksuālo) ciklu raksturo periodiskas izmaiņas dzemdes gļotādā, kas notiek saistībā ar olšūnu nobriešanas procesu olnīcā un ovulāciju.
Menstruālais cikls sastāv no diviem cikliem: olnīcu un dzemdes. Olnīcu cikls ir saistīts ar olšūnas nobriešanas procesu olnīcā. Otro ciklu sauc par dzemdes ciklu, jo visas izmaiņas, kas notiek dzemdes struktūrā un darbībā, notiek olnīcu dzimumhormonu ietekmē.
Sievietes viena menstruālā cikla laikā ir trīs dzemdes gļotādas izmaiņu fāzes: menstruālā, pēcmenstruālā un pirmsmenstruālā (4.11. att.).
Menstruālā fāze (endometrija deskvamācijas fāze) rodas, ja olšūna nav apaugļota. Deskvamācijas fāzē samazinās estrogēna un progesterona ražošana dzeltenajā ķermenī. Tā rezultātā dzemdes gļotādā parādās nekrozes perēkļi - nekroze, asiņošana. Gļotādas funkcionālais slānis pazūd un sākas nākamās menstruācijas. Šis posms parasti ilgst 3-4 dienas. Menstruāciju fāzē izplūst 40–50 ml asiņu.

Rīsi. 4.11. Sievietes olnīcu-menstruālā cikla shēma:
I - menstruālā fāze; II - pēcmenstruālā fāze; III - pirmsmenstruālā fāze: 1 - pirmatnējais folikuls olnīcā; 2 - primārie (augošie) folikuli; 3 - primārais folikuls (vezikulu grafiki); 4 - ovulācija; 5 - menstruālais dzeltenais ķermenis ziedēšanas stadijā; 6 - dzeltenā ķermeņa apgrieztā attīstība; 7 - hipofīzes priekšējā daiva; 8 - hipofīzes aizmugurējā daiva; 9 - grūtniecības dzeltenais ķermenis; 10 - apaugļošana; 11 - implantēts embrijs. FSH - follitropīna ietekme uz folikulām; LH - lutropīna ietekme uz ovulāciju un dzeltenā ķermeņa veidošanos; LTG - prolaktīna iedarbība uz dzelteno ķermeni; E - estrogēna ietekme uz dzemdi (endometrija augšana); Pg ir progesterona ietekme uz endometriju.
Pēcmenstruālā fāze (proliferācijas fāze) rodas pēc menstruācijām un ilgst 10-12 dienas. Šī fāze ir cieši saistīta ar ietekmi uz dzemdes gļotādu. estrogēns - hormoni, kas veido jaunu attīstītu folikulu. Maksimālā dzemdes gļotādas augšana tiek novērota 12-14 dienā folikulu pilnīgas nobriešanas un ovulācijas laikā. Šajā periodā dzemdes gļotādas biezums ir 3-4 mm.
Premenstruālā fāze (sekrēcijas fāze) ir būtiska dzemdes sagatavošanai grūtniecībai. Ietekmēja gestagēni - olnīcu dzeltenā ķermeņa hormoni - dzemdes gļotādas dziedzeri arvien vairāk piepildās ar izdalījumiem, šūnās uzkrājas barības vielas, palielinās savīto asinsvadu skaits. Šajā laikā dzemdes gļotāda un sievietes ķermenis ir sagatavoti apaugļotas olšūnas pieņemšanai un implantācijai.
Grūtniecības laikā palielinās dzemdes izmērs, mainās tās forma miometrija hipertrofijas dēļ. Pēc dzemdībām dzemde iegūst savu parasto formu un izmēru.
Menstruālo funkciju regulē nervu, humorālo un dzimumorgānu kompleksa kompleksa kopīgā darbība (smadzeņu garoza, hipotalāms, hipofīze, olnīcas, maksts, dzemde, olvadi).
Līdzīga informācija.
Dzemdes forma ir bumbierveida, saplacināta anteroposterior virzienā. No dzemdes augšējām sānu malām atkāpjas platas dzemdes saites, kurās atrodas dzemdes (olvados) un olnīcas (1. att.). Anatomiski dzemde ir sadalīta dibenā, ķermenī un dzemdes kaklā.
Apakšdaļa ir dzemdes daļa, kas atrodas virs olvados. Ķermenim ir trīsstūrveida forma, kas sašaurinās uz šauruma pusi. Dzemdes dobumam ir arī trīsstūra forma, augšējos stūros ir divas atveres, kas atveras olvados, apakšējā stūrī ir šaurums, kas ved uz dzemdes kakla kanāla dobumu (2. att.).
Lai izvairītos no dzirdes, var izmantot arī stetoskopu. Urologs ir kaut kas līdzīgs vīriešu ginekologam, izņemot nieru un urīnceļu ķirurģiju. Medicīnā nav nevienas operācijas, kas to darītu cietai cilvēcei kā prostatektomija. Katrs nevajadzīgs vārds ir bezjēdzīgs.
Lielie mīl, mazie tikai vairojas. Sieviete mīl vīrieti, nevis to, kāds viņš ir. Impotence joprojām ir labākais kontracepcijas līdzeklis. Daba saglabā sugu, ļoti maz rūpējas par indivīdu. Daudzu sieviešu slimību cēlonis ir vīrietis.
Grūtniecība ir specifiska infekcija, ko pārnēsā putni. Dzimšanas laikā izdariet gudru izskatu. Paturiet prātā, ka jūsu seja parasti ir pirmā lieta, ko bērns redz. Mēs novecojam tik ātri, ka naktīs, kad ir klusums, mēs dzirdam, kā darbojas artērijas.
Dzemdes kakls ir salīdzinoši šaurs dzemdes apakšējais segments. Meitenēm un meitenēm tam ir koniska forma, pieaugušai sievietei tā ir cilindriska. Izšķir maksts daļu (portio vaginalis cervicis - ectocervix), dzemdes kakla kanālu (canalis cervicalis uteri - endocervix) un šauru. Dzemdes kaklā ir divas atveres: iekšējā os - atvere augšējā daļā, kas atrodas uz ķermeņa un dzemdes kakla robežas, un ārējā os - atvere apakšējā daļā, kas atveras makstī.
Sirds ir kā nesildītas plūmju klimpas. Kardiologs, viņam nav sirds, viņš vēlas nomainīt pacientus ar sūkni. Mūsdienu triloģija: aptaukošanās - diabēts - sacietēšana. Pacienti ar intravenozu barošanu vienmēr guļ koridora pretējā galā.
Paturiet prātā, ka pat baktērijas skatās uz mums no mikroskopa otras puses. Ja mēs patiešām zinātu, kā un no kā sastāv mūsu ķermenis, mēs neuzdrošināmies kustēties. Liesai ir līdzīga funkcija kā valsts notāram. Mani vienmēr ir nicinājušas militārās lietas.
Neirologa darbs ir izpētīt, vai cilvēks kādā brīdī netiks traumēts ar muskuli pavisam citā vietā, kur neviens cits, izņemot neirologu, to nesagaidītu. Kuņģa čūlas ir lipīga slimība. Tie, kuriem jau ir, to dara ar citiem.
Dzemdes kakla maksts daļa ir noapaļota, tās virsma ir gluda, un ārējā os atrodas centrā. Sievietēm, kas nav dzemdējušas, tas ir mazs, apaļš vai šķērseniski ovāls (mazas zivs mute). Pēc dzemdībām ārējā rīkle iegūst šķērsvirziena spraugas formu. Dzemdes kakla kanāls ir šaurs, vidusdaļā paplašināts. Uz priekšējās un aizmugurējās virsmas ir divas gareniskas izciļņas, no kurām leņķī stiepjas gļotādas krokas, plaukstas formas krokas. Šie veidojumi piešķir kanālam dīvainu izskatu un tiek saukti par dzīvības koku.
Smadzenītes - smadzenītes
Tas, kurš moka acis, vairs neinteresē par angļu flotes nākotni. Starp citu, vai tu tici vidusauss? Fiziologs paskaidros, kāpēc vardei nav vajadzīgs auglis, lai gan lielākajai daļai cilvēku tas ir vajadzīgs. Vai mazohisms ir nespēja izjust baudu, kas nav sāpes, vai spēja izbaudīt sāpes?
Smadzenes ir aparāts, ar kuru mēs domājam to, ko domājam. Kosmētika nodarbojas ar ādas kopšanu, līdz tajā ienāk ādas kopšanas pakalpojumu sniedzējs. Definīcijas un mērķi, pozīcija bioloģijas zinātņu sistēmā, mācību un pētniecības nozīme farmācijas nozarēs, šūna kā ķermeņa pamatvienība, šūnu populācija - audu-orgānu sistēmas. Audi: to attīstība un diferenciācija ontoģenēzē, tipoloģijas pamati - epitēlijs un dziedzeri, saistaudi un trofiskie audi, muskuļu audi, nervu audi. Anatomiskā nomenklatūra, topogrāfiskās attiecības uz ķermeņa, kustību sistēma, vispārīgā zinātne par kauliem un muskuļiem, to savienojumu un mehāniku, cilvēka skelets, muskuļu grupu topogrāfija. Asinsvadu sistēma. Nieru un urīna aizplūšana, virsnieru dziedzeri, vīriešu reproduktīvā struktūra, sieviešu dzimumorgānu struktūra, apaugļošanās un dzimumšūnu veidošanās, diferenciācijas un noteikšanas mehānismi, kritiskie un jutīgie attīstības periodi - nozīme teratoloģijā, fetoplacentārā vienība. Nervu sistēma ir centrālā nervu sistēma. Mugurkaula nervu uzbūve un gaita, veģetatīvo nervu uzbūve un norise, redzes aparāta uzbūve un neironu ceļi, dzirdes sistēmas uzbūve un nervu ceļi, ādas un infernālā uzbūve. Definīcija, fizioloģijas saturs, homeostāze, cilvēka ķermeņa sastāvs, membrānu transports, asins sastāvs, to īpašības, asins grupas, plazma, asins koagulācija, limfa. Nieru asiņošana, nefronu glomerulārās un tubulārās funkcijas, urīnceļu darbība, urīna sastāvs un īpašības, nieru darbības regulēšana, nieru funkcionālie testi. Endokrīno dziedzeru fizioloģija. Vīriešu un sieviešu reproduktīvā sistēma, hormonālā regulācija, grūtniecība, asinsrite un augļa elpošana, dzemdības, laktācija. Gremošanas sistēmas fizioloģija. Gremošanas trakta, aknu, aizkuņģa dziedzera, žultspūšļa fizioloģija, gremošana un barības vielu uzsūkšanās. Olbaltumvielas, lipīdi, ogļhidrāti, vitamīni, minerālvielas uzturā, racionāls uzturs. Maņu orgānu fizioloģija. Vizuāli dzirdes ožas garšas sistēma, neiromuskulārā transmisija, muskuļu kontrakcija, muskuļu darbs, muskuļu aktivitātes elektrofizioloģija. Centrālās un perifērās nervu sistēmas fizioloģija. Reflekss, receptoru un nervu šķiedras, sinapse, veģetatīvā nervu sistēma, mugurkaula muguras smadzeņu darbība, muguras smadzeņu muguras smadzenes, tilts, smadzenītes, vidussmadzenes, vidussmadzenes, telencefalons.
- Citoloģija un vispārējā histoloģija.
- Orgānu sistēmu morfoloģija.
- Asinsrites sistēmas fizioloģija.
- Plaušu ventilācija, difūzija, perfūzija, elpošanas mehānisms, elpošanas kontrole.
- Ekskrēcijas sistēmas fizioloģija.
Maksts ir muskuļu elastīga caurule, kas atrodas mazajā iegurnī, augšējā daļa aptver dzemdes kaklu, bet apakšējā atveras dzimumorgānu spraugā.
Dzemdes ķermeņa gļotāda sastāv no stromas un viena slāņa cilindriska epitēlija, kas izaug stromā, veidojot vienkāršus cauruļveida dziedzerus. Straujuma gļotāda ir līdzīga dzemdes ķermeņa gļotādai, un to attēlo liels skaits saistaudu šūnu un atsevišķi vienkārši nesazarojoši dziedzeri. Menstruālā cikla laikā ķermeņa gļotādā un zarnās notiek cikliskas izmaiņas.
Noskatieties filmu: Cik bieži mēs nodarbojamies ar seksu?
Vīriešu reproduktīvā sistēma sastāv no iekšējiem orgāniem, t.i. sēklinieki, epididīms, sēklas pūslīši, folikulu epitēlija dziedzeri, ejakulācijas kanāls, prostatas un urīnizvadkanāla cauruļveida dziedzeri. Ārējie orgāni ietver sēklinieku maisiņu un dzimumlocekli.
Vīriešu dzimumorgānu uzbūve
Kodols atrodas sēkliniekos. Ārēji to pārklāj seroza membrāna un iekšēji epitēlija membrāna, kas nonāk starpsienā, kas atdala kodolus no sēkliniekiem. Tieši šajās daivās kodols atrodas kodolā. Sākumā tie ir sajaukti, bet kodola nišas zonā tie nonāk tieši kanālos un nonāk epididimālās kanāliņos.Dzemdes kakla siena sastāv galvenokārt no kolagēna audiem, gļotādas stromā ir daudz elastīgu šķiedru. Endokerviksa dziedzeri ir cauruļveida, sazaroti, tos neuzskata par īstiem dziedzeriem, jo to struktūra ir vienāda. Dziedzeri satur noslēpumu biezu stiklveida gļotu veidā, kam ir sārmaina reakcija. Sārmainā reakcija veicina spermatozoīdu dzīvotspējas saglabāšanu, to pārvietošanos dzemdes dobumā. Ovulācijas laikā palielinās gļotu sekrēcija, noslēpums aizpilda dzemdes kakla kanālu un veido tā saukto Kristellera korķi, kas, pateicoties savām baktericīdajām īpašībām, mehāniski neļauj mikrobiem iekļūt kanālā un dzemdes dobumā. Ja dziedzeri tiek bloķēti un gļotas turpina uzkrāties, veidojas nabotiešu cistas, kas var izvirzīties uz dzemdes kakla virsmas.
Starp kanāliņiem atrodas šūnas, kas ir atbildīgas par vīriešu dzimuma hormonu ražošanu. Kodolā atrodas spermatogēnais epitēlijs, kas sastāv no spermatīdiem un spermatogoniju - vīriešu reproduktīvo šūnu - spermas. Nervi pielīp pie sēkliniekiem gar muguru. Nekroze ir kanāliņi, kas veido vairākus metrus garu auklu, kurā ir skropstas, kas ir atbildīgas par spermas kustīgumu. Baro spermas krātuvi, līdz tā sasniedz pilnīgu briedumu.
No turienes spermatozoīdi nonāk iegurnī, un ārpus urīnpūšļa nonāk stomatālajā kanālā, kur tie savienojas ar sēklas pūslīšu caurulīti un veido izsviedes kanālu. Tas atrodas netālu no urīnpūšļa dibena un tiek izmantots, lai ražotu vielas, kas nodrošina enerģiju spermai.
Kanāla gļotādu attēlo cilindrisks gļotu veidojošs epitēlijs, ir atsevišķas ciliētas šūnas, to skaits ievērojami samazinās līdz ar vecumu.
Ektocerviksa un maksts gļotāda ir izklāta ar stratificētu plakanu, nekeratinizētu epitēliju. Parasti reproduktīvajā vecumā epitēlijs sastāv no daudzām rindām, kas nosacīti sadalītas trīs slāņos: bazālā, vidējā, virspusējā. Ar bazālo membrānu ir savienots tikai apakšējais (bazālais) šūnu slānis, tajā esošās šūnas ir sakārtotas vienā rindā. Jaunu šūnu slānis, kas atrodas virs bazālā slāņa, kas sastāv no vairākām rindām (starpslāņa apakšējā daļa). To sauc par parabazālu. Šūnu izmērs palielinās līdz ar briedumu. Kodolu izmērs samazinās (3. att.)
Ejakulācijas caurule un prostatas dziedzeris
Injekcijas caurule atrodas prostatas iekšpusē. Tā platums sašaurinās urīnizvadkanāla mutē. Sīpolu un cauruļveida dziedzeri ir atbildīgi par precum sekrēciju no urīnizvadkanāla, noslēpuma, kas aizsargā spermu no urīnizvadkanāla un maksts skābās vides. Šī ir ādas soma, kas atrodas vulvas zonā. Sēkliniekos ir sēklinieki.
Dzimumloceklis apvieno vīriešu reproduktīvās sistēmas un urīnceļu funkcijas. To lieto gan urīna izvadīšanai no urīnpūšļa, gan spermas ievadīšanai sievietes dzimumorgānos, kas ir dzimumlocekļa šķīdums. Āda, kas pārklāj dzimumlocekli, ir plāna un caurspīdīga, savukārt priekšāda ir priekšāda.
Maksts un dzemdes kakla maksts daļas slāņveida plakanā epitēlija struktūra ir atkarīga no sievietes hormonālā stāvokļa, menstruālā cikla fāzes. Menstruālā cikla laikā ir 4 fāzes: menstruālā, folikulīna (estrogēna, proliferatīvā), ovulācijas un luteālā (progestīna, sekrēcijas). Šīs fāzes ir saistītas ar olšūnas nobriešanu, ko regulē hipotalāma-hipofīzes sistēmas (hipofīzes priekšējās daļas) gonadotropie hormoni. FSH ietekmē folikuls aug un nobriest olnīcā (4. att.).
Seksuālajai sistēmai ir jāveic ārkārtīgi svarīgi uzdevumi. Viņš ir atbildīgs ne tikai par sieviešu hormonu ražošanu, bet arī par olšūnu veidošanos, par augļa attīstību un pārtraukumu. Viņa lielās atbildības dēļ nav pārsteidzoši, ka sievietes seksuālā sistēma dažreiz sāk trīcēt.
Peles - struktūra, funkcijas un slimības
Pelēm ir svarīga loma sieviešu reproduktīvās sistēmas darbībā. Tas ir tikai viņas iekšienē, tā ir olšūna, olšūna, ko viņas dzemde aizsargā un baro nākamos 9 mēnešus. Tie nodrošina arī svarīgas nereproduktīvās funkcijas, īpaši kaulos, muskuļos, ādā, hematopoētiskajā sistēmā, centrālajā nervu sistēmā. Androgēnu ražošanas vieta ir gan vīriešu, gan sieviešu dzimumdziedzeri, virsnieru dziedzeri un vēdera garoza. Androgēni ir steroīdi, atvasinājumi.
Augošais folikuls izdala estrogēnu hormonus, kuru noteikts daudzums kavē FSH veidošanos un stimulē LH izdalīšanos. LH kopā ar FSH sagatavo folikulu ovulācijai, un pēc folikula plīsuma (ovulācijas), ja grūtniecība nav iestājusies, veicina tā pārvēršanos dzeltenajā ķermenī.

Tāpat ir nepareizi sievietes veselību saistīt tikai ar viņas reproduktīvo funkciju, kas liek meklēt visu patoloģiju cēloņus tikai tad, kad mainās reproduktīvā sistēma. Šobrīd viņš cīnās ar šo pārliecību. Lēna attīstība t.s. seksa zāles.
Viņu darbība neaprobežojas tikai ar reproduktīvās sistēmas darbību. No pirmajiem līdz pēdējiem mēnešiem tie pasargā sievietes sirdi un asinsvadus no meningokoku infekcijas. Nodrošina ādas un gļotādu mitrināšanu. Vai urologs. Ja jums ir iespēja ārstēties profesionālā neauglības klīnikā, varat to izmantot. Visi konsultanti, kā arī profesionāla laboratorija.
Maksts epitēlijs ir visvairāk jutīgs pret hormonālo ietekmi, tāpēc hormonālās citoloģiskās diagnostikas pamatā ir šī epitēlija sastāva izpēte (sk. Hormonālā citoloģiskā diagnostika).
Kanāla cilindriskais epitēlijs pāriet plakanā tā sauktajā krustojuma zonā, kas atrodas meitenei dzemdes kakla maksts daļā, sievietei reproduktīvā vecumā - ārējās os līmenī. Hormonālu un citu ietekmju ietekmē locītavu zona var pārvietoties uz dzemdes kakla maksts daļu. Cilindriska epitēlija parādīšanos uz kakla maksts daļas sauc par ektopiju. Maksts satura ietekmē ektopijas vietā notiek fizioloģiskas izmaiņas, metaplāzija par plakanu epitēliju (6. att.).
 |
1 - nobriedis epitēlijs. Uz virsmas nobriedušas virsmas slāņa šūnas ar piknotiskiem kodoliem 2 - epitēlijs nobriest līdz starpslānim, nobriedušas starpposma šūnas uz virsmas 3 - epitēlijs nobriest līdz starpslānim, uz virsmas nenobriedušas starpšūnas Statīvs neko nesavieno. Tā var nebūt sēnīšu infekcija, un jums būs jāmaina medikamenti, piemēram, antibiotikas. Asiņošana no reproduktīvās sistēmas ir ļaundabīga. Kad esat veicis kādu pētījumu, varat to izslēgt, ja asiņošana notiek pēc katrām attiecībām un nav tendence apstāties. Ieteicams apmeklēt ginekologu, veikt laboratorijas pārbaudi. Vai neatradāt to, ko meklējāt? Mātītes reproduktīvie orgāni atrodas iegurnī, un tie ietver olnīcas, olšūnas, dzemdi un maksts. Tie ir parādīti zemāk esošajā attēlā. Anteroposterior skats parāda sieviešu reproduktīvās sistēmas diagrammu. Skats no sāniem, kas parāda sievietes reproduktīvo orgānu stāvokli attiecībā pret urīnpūsli un taisno zarnu. 4 - 5 - epitēlijs nobriest tikai līdz parabazālajam slānim, parabazālās šūnas uz virsmas |
5. att. Dažādas slāņveida plakanšūnu epitēlija nobriešanas stadijas |
|
 |
A - pirms pubertātes (savienojuma zona atrodas dzemdes kakla kanālā) B - C - pubertātes laikā (savienojuma zona tiek pārvietota uz dzemdes kakla maksts daļu) Sieviešu reproduktīvā trakta shēma. Sieviešu reproduktīvā trakta šķērsgriezums, kas ilustrē urīnpūšļa telpisko un taisnās zarnas savienojumu. Olas ir gluds orgāns. Tie atrodas abās dzemdes pusēs, blakus iegurņa sānu sienai. Olas ir atbildīgas par oģenēzi un ciklisku ovulāciju sievietēm pēc pubertātes. Olnīcām ir endokrīnā iedarbība, kas sastāv no hormonu estrogēna un progesterona ražošanas, kas kontrolē folikulu nobriešanu un sagatavo dzemdi apaugļotas olšūnas saņemšanai. Olu un saistīto hormonu attīstības stadiju kopsavilkums. Mātītes ir divi apmēram 10 cm gari vadi, pa kuriem olšūna tiek pārnesta no olnīcas virsmas uz dzemdes dobumu. Piltuvveida olvadu mute ieskauj olnīcas virsmu un spēlē lomu olšūnas kustībā uz olvadu. Olšūna tiek transportēta pa olvadu, veicot maigām peristaltiskām olvadu kustībām un cauruļveida epitēlija viena slāņa epitēlija rullīti, kas izklāj olvadu. Olvada gļotāda ir saburzīta, kas nodrošina pareizos apstākļus olšūnas apaugļošanai. D - maksts daļas cilindriskais epitēlijs tiek aizstāts ar metaplastisku D - dzemdes kakla kolposkopisks attēls daudzdzemdējušai sievietei |
6. att. Kolonnveida epitēlija un savienojuma vietas atrašanās vieta dzemdes kaklā |
|
Hipofīzes LH ietekmē izveidojies dzeltenais ķermenis atbrīvo progesteronu. Pastāv cieša saikne starp olnīcu un virsnieru dziedzera, olnīcu un vairogdziedzera hormoniem.
Estrogēni stimulē noslāņotā plakanā nekeratinizētā epitēlija pilnīgu nobriešanu līdz virsmas šūnām. Progesterons kavē nobriešanu, un, ja tas tiek ražots lielos daudzumos, šūnas nobriest tikai līdz starpslānim. Pēcmenopauzes periodā dzimumhormonu ražošanas samazināšanās dēļ epitēlijs atrofē (5. att.).
Cilindriskais epitēlijs maksts daļā tiek aizstāts ar metaplastisku. Metaplastiskā epitēlija laukumu sauc par transformācijas zonu vai transformācijas zonu. Sievietēm reproduktīvā vecumā savienojuma zonu parasti attēlo dabiskā savienojuma zonas zonas, transformācijas zonas zonas un metaplastiskais epitēlijs. Transformācijas zona var atrasties dzemdes kakla maksts daļā, vai arī tā var (pilnībā vai daļēji) nonākt dzemdes kakla kanālā. Sievietēm pēcmenopauzes periodā krustojuma zona un transformācijas zona visbiežāk atrodas dzemdes kakla kanālā. Transformācijas zona ir visbīstamākā patoloģisku, tostarp neoplastisku izmaiņu iespējamības ziņā.
Metaplastiskais plakanšūnu epitēlijs (7. att.) veidojas nevis no nobriedušām cilindriskām, bet no subcilindriskām, tā sauktajām rezerves šūnām. Parasti rezerves šūnas parasti nav atrodamas histoloģiskajos un citoloģiskajos preparātos. Rezerves šūnu hiperplāzija ir pirmā plakanšūnu metaplāzijas stadija. Zem cilindrisko šūnu slāņa parādās viens, divi vai vairāki germinālā tipa šūnu slāņi, kas līdzinās plakanā epitēlija bazālā slāņa šūnām bez skaidrām šūnu robežām.
Nobriedis metaplastiskais epitēlijs morfoloģiski praktiski neatšķiras no "dabiskā" plakanā epitēlija, ko pārstāv visi slāņi, kas raksturīgi stratificētam plakanam nekeratinizētam epitēlijam.
 |
A - kolonnu epitēlijs B - zem cilindriskā epitēlija slāņa parādās subcilindrisku (rezerves) šūnu slānis B - rezerves šūnas vairojas, cilindriskās tiek noslīdētas no virsmas D - nenobriedušas plakanās metaplāzijas stadija: tiek noteiktas skaidras rezerves šūnu robežas un pakāpeniski veidojas 3-4 šūnu slāņi, līdzīgi kā stratificēts plakanšūnu nekeratinizēts epitēlijs. D - plakanšūnu metaplāzijas nobriešanas stadija. Uz epitēlija slāņa virsmas vidēja izmēra šūnas ar maziem kodoliem E - nobriedušas plakanšūnu metaplāzijas stadija. Šūnas uz epitēlija slāņa virsmas ir līdzīgas plakanā epitēlija starpšūnām G - nobriedušas plakanšūnu metaplāzijas stadija. Šūnas uz epitēlija slāņa virsmas praktiski neatšķiras no plakanā epitēlija virsmas slāņa "dabiskajām" šūnām. |
7. att. Plakanās metaplāzijas stadijas |
|
Tatarčuks T.F., Solsky Ya.P., Regeda S.I., Bodrjagova O.I.
1. attēls. Reproduktīvās sistēmas funkcionālā struktūra
Lai pareizi klīniski novērtētu sievietes ķermeņa neiroendokrīnos traucējumus un attiecīgi noteiktu to patoģenētiskās terapijas principus un metodes, pirmkārt, ir jāzina reproduktīvās sistēmas piecu saišu regulējums, galvenā funkcija. no kuriem ir bioloģiskas sugas vairošanās (1. att.).
1. nodaļa. Sieviešu reproduktīvās sistēmas anatomija un fizioloģija 9
Reproduktīvās sistēmas funkciju regulēšanu nosaka hipotalāma-hipofīzes saite, kuru, savukārt, kontrolē smadzeņu garoza caur neirotransmiteriem un neirotransmiteriem (Lakoski J.M., 1989).
Hipotalāms ir sava veida ķermeņa bioloģiskais pulkstenis, tas ir, neiroregulācijas procesu pašregulācijas un automatizācijas sistēma, kas īsteno informāciju, kas nāk no ķermeņa ārējās un iekšējās vides, tādējādi nodrošinot iekšējo homeostāzi, kas nepieciešama normālai ķermeņa gaitai. fizioloģiskie procesi. Tieši hipotalāms ir galvenā saite, kas koordinē hipotalāma-hipofīzes-olnīcu kompleksa darbību, kura darbību regulē gan CNS neiropeptīdi, gan olnīcu steroīdi ar atgriezeniskās saites mehānismu (Wildt L., 1989; Sopelak V.M., 1997). ).
Ņemot vērā diezgan labu reproduktīvās sistēmas perifērās daļas atspoguļojumu mūsdienu literatūrā, kā arī arvien pieaugošā psihoemocionālā stresa pieaugošo lomu dishormonālo traucējumu attīstības mehānismos, mēs uzskatījām par lietderīgu sīkāk pakavēties pie šī jautājuma. daži suprahipotalāmu struktūru līdzdalības aspekti reproduktīvās sistēmas regulēšanā.
Kā zināms, smadzenes sastāv no divu veidu šūnām: neironiem, kas veido 10% no visām smadzeņu šūnām, un glia – astrocītiem un oligodendrītiem, kas veido attiecīgi atlikušos 90%.
Neironu un glia attīstība notiek no neiroepitēlija prekursora - cilmes šūnas, kā rezultātā notiek 2 šūnu līniju sintēze: neironu prekursoru šūnas, no kurām rodas dažāda veida neironi, un glia prekursoru šūnas, no kurām pēc tam attīstās astrocīti un oligodendro. -cytes (Lakoski J.M., 1989; Sopelak V.M., 1997).
Neironi ir ļoti diferencētas šūnas ar atšķirīgiem izmēriem, formām un intracelulārām organellām. Tāpat kā visām pārējām šūnām, izņemot eritrocītus, neironiem ir šūnas ķermenis, kura centrā atrodas kodols, ko ieskauj dažāda tilpuma citoplazma.
No neironu virsmas atzarojas uztveršanas procesi - dendriti un vienīgais galvenais pārraides process - aksons, kas sniedzas līdz tā specifiskajām sinaptiskajām mērķa šūnām un var ievērojami atšķirties garumā (Sopelak V.M., 1997).
Neirona galvenais dzīves process koncentrējas šūnas ķermeņa citoplazmā (saukts arī par perikarionu), un pēc tam neironu sintēzes produkti tiek transportēti uz aksoniem un dendritiem. Divpusējs transports starp šūnas ķermeņa daļām un distālajiem procesiem nodrošina neironu funkcijas integritāti un ir pastāvīgs no enerģijas atkarīgs labi koordinēts process.
10 Endokrīnā ginekoloģija
Gliālās šūnas (no angļu valodas vārda glue — līme) sākotnēji tika uzskatītas par atbalsta smadzeņu šūnām, taču jaunākie pētījumi atklāja to svarīgo funkcionālo lomu neironu dzīves regulēšanā. Šī neneironālo šūnu elementu klase, kas ir 9 reizes lielāka par neironu skaitu, faktiski nodrošina mijiedarbību starp tiem.
Daudzskaitlīgākās glia šūnas tiek sauktas par astrocītiem to daudzizvirzīto kontūru dēļ. Šīm šūnām ir raksturīga unikāla glia fibrilārā skābā proteīna ekspresija, un tās atrodas starp asinsvadu, neironu un to savienojumu ārējo virsmu (2. att.). Astrocītu procesi tiek virzīti no neironiem uz kapilāriem, kur tie veido perivaskulāru bāzi.
2. attēls. Neironu, astrocītu un oligodendrocītu attiecības (Yen S.S.C., 1999)
Astrocītu kapilārā bāze aptver apmēram 85% no cilvēka smadzeņu kapilāriem un veido hematoencefālisko barjeru.

1. nodaļa. Sieviešu reproduktīvās sistēmas anatomija un fizioloģija 11
Vēl viena svarīga glia šūnu klase ir oligodendrocīti (šūnas ar dažiem īsiem un bieziem procesiem), kas veido aksonu mielīna apvalku, kas ļauj neironiem ātri un bez novājināšanās iedarboties lielos attālumos nervu sistēmā. Oligodendrocīti satur arī P450 steroīdu ģenēzes enzīmus un ražo pregnanolonu no holesterīna.
Steroidoģenēzes enzīmu noteikšana smadzeņu audos bija viens no atklājumiem, kas palīdzēja atklāt CNS līdzdalības mehānismus reproduktīvās funkcijas regulēšanā un, kas ne mazāk svarīgi, izskaidro CNS izmaiņas hormonālās homeostāzes izmaiņu ietekmē.
Neiroaktīvo steroīdu sekrēcija astrocītos ir augstāka nekā oligodendrocītos un neironos, un tāpēc ir nepieciešams sīkāk pakavēties pie šo šūnu īpašībām.
Astrocītu īpašības ir atšķirīgas un vēl nav pilnībā izprastas, lai gan jau ir pierādījumi, ka astrocīti ir neironu parakrīnas šūnas:
Astrocītos atklājās insulīnam līdzīgā augšanas faktora (IGF) klātbūtne, kura saturs palielinās pubertātes periodā, kā arī palielinās estrogēnu terapijas laikā;
Pituicīti kā astrocītu veids ir galvenie neironu šūnu elementi neirohipofīzē, un tiem ir svarīga loma oksitocīna un vazopresīna izdalīšanās kontrolē no neirosekrēcijas nervu galiem;
Luteinizējošā hormona (LH) un cilvēka horiona gonadotropīna (HCG) receptoru klātbūtne astrocītos liecina, ka LH un CG var ietekmēt glia šūnu darbību un attiecīgi arī smadzeņu attīstības procesus un darbību;
Astrocīti spēj ražot dažādas imūnmodulējošas molekulas, piemēram, interleikīnus (IL-1, IL-2, IL-6), audzēja nekrozes faktoru a, transformējot augšanas faktoru-os, interferonu un prostaglandīnu E, savukārt prolaktīns inducē mitoģenēzi un citokīnu ekspresiju. astrocītos;
Astrocīti, tāpat kā neironi, spēj ražot kortikotropīnu atbrīvojošo faktoru saistošo proteīnu (CRF-BP), kas ir plaši izplatīts smadzenēs. Steroīdi, piemēram, deksametazons, hidrokortizons un mazākā mērā dehidroepiandrosterons kavē CRF-SP izdalīšanos no astrocītiem;
Hipotalāma izcelsmes astrocīti izdala transformējošo augšanas faktoru a un (3), kas stimulē gonadotropīnu atbrīvojošo hormonu (Gn-RH) gēnu ekspresiju neironos, savukārt hipotalāma astrocīti ir aptuveni 4 reizes aktīvāki nekā kortikālie astrocīti attiecībā uz šūnu sintēzi. dehidroepiandrosterons (DHEA).
Astrocīti var būt iesaistīti arī glutamāta neirotransmitera līmeņa regulēšanā, kas nodrošina ierosinošu efektu, un γ-aminosviestskābes (GABA), kam ir galvenā loma anksiolītiskā (nomierinoša) efekta sasniegšanā.
12 Endokrīnā ginekoloģija
Pašlaik ir identificētas 3 galvenās raidītāju ķīmiskās formas: aminoskābes, monoamīni un neiropeptīdi.
Aminoskābes darbojas kā raidītāji gan uzbudinoši, gan nomācoši. Raidītāju vielu ierosinošajos savienojumos galvenais ir acetilholīns, kā arī glutamāts un aspartāts. Inhibējošos savienojumus regulē aminoskābes, piemēram, GABA un glicīns.
Monoamīni kā tulkotāji sastāv no kateholamīnerģiskajiem (epinefrīna, norepinefrīna un dopamīna) un serotonīnerģiskajiem raidītājiem. Tādējādi tirozīns no asinsrites nonāk kateholamīna neironos un ir substrāts, no kura tirozīna hidroksilāze katalizē dopa sintēzi. Aminoskābes dekarboksilāze (AKD) pārvērš dopa par dopamīnu. Dopamīns-(3 oksidāze (DVO) noradrenerģiskajos neironos pārvērš dopamīnu par norepinefrīnu (NA).
DA un NA izdalās sinaptiskajā spraugā, kur tie ātri saistās ar postsinaptiskajiem receptoriem. Plazmā liekos raidītājus vai nu metaboliski inaktivē katehol-O-metiltransferāze (COMT), vai atpakaļsaiste ar presinapsiskajiem receptoriem, kur tie metaboliski sadalās ar monoamīnoksidāzes (MAO), veidojot dehidroksifeniletilglikolu (DOPEG).
Peptīdu raidītāji. Hipotalāma peptīdus saturošie neironi sākotnēji tika aprakstīti kā neirosekrēcijas neironi, bet vēlāk kļuva zināms, ka gandrīz visi hipotalāma neiropeptīdi projicē daudzus smadzeņu apgabalus. Tie nodrošina neirotransmitera funkcijas pārtikas uzņemšanas, ēšanas un seksuālās uzvedības regulēšanā (1. tabula).
Atsevišķi jāpakavējas pie slāpekļa oksīda lomas centrālajā un perifērajā nervu sistēmā, kuras atklāšana radikāli mainīja iepriekš pastāvošos uzskatus par sinaptisko transmisiju. Lai gan ir būtiski pierādījumi, ka slāpekļa oksīds darbojas kā neirotransmiters, jāņem vērā, ka tas ir neparasts raidītājs, jo tā ir labila gāze, ko nevar uzglabāt sinaptiskos pūslīšos. Slāpekļa oksīds tiek sintezēts no L-arginīna ar oksidazot sintetāzes palīdzību un no nervu galiem nonāk vienkāršas difūzijas ceļā, nevis eksocitozes ceļā kā citi neirotransmiteri (3. att.). Turklāt slāpekļa oksīds neiziet atgriezeniskas reakcijas ar receptoriem, tāpat kā visi citi atgriezeniskie neirotransmiteri, bet veido kovalentus savienojumus ar vairākiem iespējamiem mērķiem, kas ietver tādus enzīmus kā izonilāta ciklāze un citas molekulas.
: atgriezenisko neirotransmiteru darbību ierobežo presinaptiskā izdalīšanās vai enzīmu degradācija, savukārt slāpekļa oksīda darbību nodrošina difūzija prom no mērķiem vai kovalento saišu veidošanās ar superoksīda anjonu.
Slāpekļa oksīda veidošanos no arginīna smadzenēs katalizē oksidazota sintetāze skābekļa klātbūtnē ar NADP kā koenzīmu
1. nodaļa. Sieviešu reproduktīvās sistēmas anatomija un fizioloģija 13
1. tabula Peptīdu raidītāji CNS
(saskaņā ar Yen S.S.C., 1999 ar izmaiņām un papildinājumiem)

un tetrahidrobioproteīns kā kofaktors. Runājot par slāpekļa oksīda lomu reproduktīvās sistēmas centrālajā regulēšanā, jāatzīmē, ka NO ir neirotransmiters, kas regulē GnRH izdalīšanos.
Neirosteroīdi. Vietējās estrogēnu sintēzes atklāšana hipotalāmā (Naftollin et al, 1975) liecināja, ka smadzenēm ir steroidoģenēzes funkcija. 1981. gadā pieaugušu žurku tēviņu smadzenēs tika konstatēta pregnanolona un pregnanolona sulfāta, kā arī dehidroepiandrosterona (DHEA) un dehidroepiandrosterona sulfāta (DHEA-S) klātbūtne. Tas noveda pie steroīdu biosintēzes mehānismu atklāšanas CNS, ko sauc par neirosteroīdiem.
Cilvēka smadzenēs neirosteroīdi, tāpat kā neirotransmiteri, ir sastopami vīriešiem un sievietēm, kas vecāki par 60 gadiem. DHEA, pregnanolons un progesterons atrodas visās smadzeņu daļās, un to koncentrācija smadzenēs ir vairākas reizes augstāka nekā plazmā.
Smadzenēs tika atklāta arī DHEA sulfāta transferāzes un sulfāta tase klātbūtne, tāpēc var pieņemt, ka DHEA-S sintēze notiek tieši smadzenēs.
14 Endokrīnā ginekoloģija

H attēls. Slāpekļa oksīda veidošanās smadzenēs (Yen S.S.C., 1999)
Steroidogēnais faktors-1 (SF-1), audu specifisks kodolreceptors, regulē vairāku steroidoģenēzes enzīmu gēnus un ir plaši sastopams cilvēka smadzenēs, tostarp limbiskās sistēmas komponentos.
Neirosteroīdiem ir ārkārtīgi svarīga loma visos organisma dzīvības procesos, tie modulē GABA receptoru, glutamāta receptoru aktivitāti, ietekmē kognitīvās funkcijas, trofiski iedarbojas uz nervu audiem (veicina mielinizāciju), modulē atbrīvojošo hormonu veidošanos organismā. hipotalāmu (Yen S.S.C., 1999).

4. attēls. Hipotalāma-hipofīzes savienojuma sagitālais griezums (Solepak V.M., 1997)
Hipotalāms ir diencefalona daļa, kas atrodas zem trešā kambara starp optisko chiasmu un vidējo eminenci, kas caur hipofīzes stumbru savienojas ar hipofīzes mugurējo daļu, kā arī savienojas ar pārī savienotajiem mastoīdajiem ķermeņiem (4. att.).
1. nodaļa. Sieviešu reproduktīvās sistēmas anatomija un fizioloģija 15
Hipotalāms ir savstarpēji saistīts ar CNS un hipofīzi, izmantojot dažādus asinsrites un nervu savienojumus. Tas sastāv no nervu šūnām, kas sagrupētas kodolos. Šūnas, kas sagrupētas hipotalāma paraventrikulārajos un supraoptiskajos kodolos, turpinās uz aizmugures hipofīzi, kur izdalās vazopresīns, oksitocīns un neirofizīni. Tajā pašā laikā supraoptiskajiem un paraventrikulāriem kodoliem ir tieša neironu saikne ar hipofīzes aizmugurējo daļu. Supraoptiskie kodoli galvenokārt izdala vazopresīnu, bet paraventrikulārie kodoli izdala oksitocīnu, kas tiek transportēts pa nervu galiem uz aizmugurējo daivu (Sopelak V.M., 1997).
Citi kodoli ražo atbrīvojošos un inhibējošos faktorus (Gn-RH, TRH, somatostatīnu, kortikotropīnu atbrīvojošo hormonu (CRH), kas tiek transportēti uz hipofīzes priekšējo daļu caur asinsrites portālu sistēmu un kontrolē hipofīzes priekšējās daļas sekrēciju.

5. attēls. Hipofīzes sagitālā daļa (Solepak V.M., 1997)
Funkcionālos savienojumus ar hipofīzes priekšējo daļu attēlo hipotalāma-hipofīzes asinsvadu sistēma (Wildt L., 1989). Hipotalāma hormoni iekļūst priekšējā daivā caur mediālo eminenci un hipotalāma-portāla cirkulāciju. Hipotalāmam ir arī intrahipotalāmu neironu savienojumi, aferento šķiedru savienojumi ar vidussmadzenēm un limbisko sistēmu, eferento šķiedru savienojumi ar vidējo smadzeņu un limbisko sistēmu, kā arī hipofīzes aizmugurējā daļa. Hipotalāma faktori tiek transportēti pa nervu šķiedrām uz vidējo eminenci, kur tie iekļūst hipofīzes kapilāru sieniņās (5. att.). Šie faktori ietekmē hipofīzes endokrīnās šūnas un nodrošina specifiskas hormonālās atbildes (Yen S.S.C., 1999).
16 Endokrīnā ginekoloģija
Runājot par reproduktīvās sistēmas regulēšanu, jāuzsver, ka hipotalāmu atbrīvojošo hormonu ietekmē tiek veikta gonadotropo hormonu sintēze hipofīzē. Hipofiziotropo atbrīvojošo hormonu (liberīnu), kas pēc ķīmiskās dabas ir dekapeptīdi, sintēzes vieta ir tieši mediobazālā hipotalāma lokveida kodoli. Atbrīvojošo hormonu ražošana notiek noteiktā pulsējošā ritmā, ko sauc par cirkorālu.
Lai nodrošinātu normālu gonadotropīnu sekrēciju, pietiek uzturēt stabilu GnRH fizioloģiskā daudzuma izdalīšanās biežumu. Izmaiņas GnRH izdalīšanās biežumā maina ne tikai hipofīzes izdalītā LH un FSH daudzumu, bet arī to attiecību, savukārt pat desmitkārtīga GnRH koncentrācijas palielināšanās izraisa tikai nelielu FSH izdalīšanās palielināšanos un neizmaina. jebkādā veidā mainīt LH sekrēciju (Halvorson L.M. et al., 1999).
Tādējādi ritma palielināšanās ievērojami palielina FSH izdalīšanos un samazina LH izdalīšanos. Luteālajā fāzē progesterons ar endogēno opiātu starpniecību palēnina impulsu ģeneratora frekvenci, un šo darbību nosaka nevis progesterona koncentrācija, bet gan tā iedarbības ilgums. Estradiols, iedarbojoties uz hipotalāmu un gonadotropiem (palielina Gn-RH receptoru blīvumu), palielina LH / FSH viļņa amplitūdu.
GnRH izdalīšanās biežums cilvēkiem ir 1 izdalīšanās 70-90 minūtēs un atbilst vairākiem bioritmiem (miega fāžu maiņa, glomerulārās filtrācijas un kuņģa sekrēcijas ātruma svārstības, karstuma viļņu biežums menopauzes laikā utt.) . Informācijas frekvences modulācija nodrošina reproduktīvās sistēmas regulēšanas ātrumu un uzticamību un izturību pret traucējumiem.
Ritma impulsa ģenerators - hipotalāma lokveida kodols fizioloģiskos apstākļos saņem informāciju par gonadotropīnu izdalīšanos no hipofīzes, izmantojot īsu atgriezeniskās saites sistēmu, jo īpašie sfinkteri regulē spiediena gradientus portāla asins plūsmas sistēmā, un daļa no asinis no hipofīzes atgriežas hipotalāmā, kas nodrošina ļoti augstu vietējo hipofīzes hormonu koncentrāciju hipotalāmā (Yen S., 1999).
LH un FSH sintēzi un sekrēciju hipofīzē veic tās pašas šūnas (Halvorson L.M. et al., 1999). Uz gonadotrofu virsmas atrodas GnRH receptori, kuru blīvums ir atkarīgs no steroīdo hormonu līmeņa asinīs un no GnRH koncentrācijas. Gn-RH kombinācija ar receptoru izraisa masīvu kalcija jonu pieplūdumu šūnā, kas pēc dažām minūtēm noved pie LH un FSH izdalīšanās asinsritē. Turklāt GnRH stimulē LH un FSH sintēzi un saglabā gonadotrofu integritāti (Wildt L., 1989).
Svarīga loma endokrīno dziedzeru darbības regulēšanā pieder hipofīzei. Tas atrodas turku seglos pie smadzeņu pamatnes, sastāv no priekšējās (adenohipofīzes), vidējās un aizmugurējās (neirohipofīzes) daivas. Starpposma daivas cilvēkiem praktiski nav. Hipofīze savienojas ar hipotalāmu caur hipofīzes stumbru (skat. 5. att.).
1. nodaļa. Sievietes reproduktīvās sistēmas anatomija un fizioloģija I?
Hipofīzes priekšējā daļa sastāv no pieciem dažādu veidu šūnām, kas atšķiras pēc imunoloģiskām un ultrastrukturālajām īpašībām. Šīs priekšējās daivas šūnas ražo 6 zināmus hormonus:
adrenokortikotropais hormons (AKTH) vai kortikotropīns;
vairogdziedzeri stimulējošais hormons (TSH) vai tirotropīns;
Gonadotropie hormoni: folikulus stimulējošais (FSH) vai folitropīns un luteinizējošs (LH) vai lutropīns;
somatotropais hormons (GH) vai augšanas hormons;
Prolaktīns.
Pirmie 4 hormoni regulē tā saukto perifēro endokrīno dziedzeru funkcijas, savukārt augšanas hormons un prolaktīns iedarbojas tieši uz mērķa audiem (Halvorson L.M. et al., 1999).
Augšanas hormonu un prolaktīnu ražo divu veidu šūnas - somatotrofi un laktotrofi (mammotrofi), kas pieder pie acidofilās sērijas. AKTH un citas proopiomelatokortīna molekulu frakcijas, piemēram, p-lipotropīnu un endorfīnus, sintezē tirotrofi, savukārt LH un FSH sintezē gonadotrofi, kas pieder pie bazofīlās sērijas.
Gonadotrofi veido 10-15% no hipofīzes priekšējās daļas šūnu sastāva un atrodas laktotrofu tuvumā. Šī lokalizācijas iezīme liecina, ka starp diviem šo šūnu veidiem pastāv parakrīnas attiecības (Sopelak V.M., 1997).
Kā jau minēts, šo sešu priekšējās daivas hormonu sekrēciju kontrolē hipotalāmu atbrīvojošie un inhibējošie faktori, kas izdalās no hipotalāma un nonāk hipofīzē caur hipotalāma-hipofīzes portāla traukiem. Taču tropisko hormonu veidošanos var ietekmēt arī citas vielas, kas sintezētas gan reproduktīvās sistēmas centrālajā (P-endorfīni), gan perifērajā (estradiols) daļās (Halvorson L.M. et al., 1999).
Neirohipofīze ietver hipofīzes stumbru (skat. 5. att.), nervu daivu un vidējo eminenci (īpaši neironu audi hipotalāma pamatnē, kas veido galveno reģionu hipofīzes regulējošo neirosekrēciju pārnešanai uz hipofīzes priekšējo daļu ). Divi aizmugures hipofīzes hormoni (vazopresīns un oksitocīns) uzkrājas granulās ar attiecīgajiem neirofizīniem, tiek transportēti pa aksoniem un samontēti aksonu terminālos, kur tie tiek uzglabāti līdz atbilstošiem impulsiem, kas izraisa to atbrīvošanos. Neiropeptīdi tiek atbrīvoti no sekrēcijas granulām eksocitozes ceļā. Šis process ietver neirosekrēcijas granulu membrānu un nelielas šūnas membrānas daļas izšķīšanu aksona galā. Granulu saturs nonāk starpšūnu telpā un no turienes asinsritē (Sopelak V.M., 1997).
Reprodukcijas regulēšanu un dzimumdziedzeru darbību galvenokārt veic adenohipofīzes izdalītie gonadotropie hormoni, proti, FSH, LH un prolaktīns. FSH - izraisa granulozes proliferāciju
Endokrīnā ginekoloģija
Šūnas, stimulē folikulu augšanu. LH – aktivizē androgēnu sintēzi un kopā ar FSH veicina ovulāciju. FSH un zāļu sekrēciju regulē gonadotropīnu atbrīvojošais hormons ar atgriezeniskās saites mehānismu, un tas ir atkarīgs arī no estrogēnu un androgēnu līmeņa. Gonadoliberīns (Luliberīns) tiek izdalīts ar impulsiem ar frekvenci no 1 impulsa stundā līdz 1-2 impulsiem dienā. Gonadoliberīna sekrēciju kontrolē dzimums un citi hormoni, daudzi CNS neirotransmiteri, tostarp kateholamīni, opiātu hormoni utt. Gonadoliberīns mijiedarbojas ar receptoriem, kas atrodas uz gonadotrofu membrānām, un, lai aktivizētu receptoru, ir nepieciešama pirmo trīs aminoskābju klātbūtne. Gonadoliberīna agonisti (buzerilīns, nafarelīns, leuprolīds utt.) iedarbojas, mijiedarbojoties ar tiem pašiem membrānas receptoriem (Halvorson L.M., 1999).
Prolaktīns kavē gonadotropo hormonu veidošanos. Glikokortikoīdiem ir arī inhibējoša iedarbība uz LH izdalīšanos.
Pēc ķīmiskās struktūras LH un FSH ir glikoproteīni, kas sastāv no divām polipeptīdu apakšvienībām a un p. šo hormonu a-apakšvienība ir kopīga katram glikoproteīnam un tai ir tāda pati aminoskābju secība, P-apakšvienība starp glikoproteīniem atšķiras ar tajā esošo aminoskābju secību. Tā ir P-apakšvienība, kas ir atbildīga par hormonālo specifiku. Abas apakšvienības ir atsevišķi bioloģiski neaktīvas. Heterodimēru veidošanās ir priekšnoteikums bioloģiskās aktivitātes izpausmei (Halvorson L.M., 1999).
Asinīs cirkulējošo gonadotropīnu pussabrukšanas periods ir tieši saistīts ar sialskābes komponentu hormona molekulā. Ir pierādīts, ka desialilācija saīsina gonadotropīnu pussabrukšanas periodu un bioloģisko aktivitāti. FSH ir asinīs brīvā formā, un tā pussabrukšanas periods ir 55-60 minūtes, bet LH ir 25-30 minūtes. Reproduktīvā vecumā LH izdalīšanās dienā ir 500-1100 mIU, pēcmenopauzes periodā LH veidošanās ātrums palielinās un tā daudzums ir līdz 3000-3500 mIU dienā (Sopelak V.M., 1997).
Tāpat kā steroīdi, gonadotropīniem ir bioloģiska ietekme uz mērķa audiem, aktivizējot specifiskus receptorus. Tomēr atšķirībā no steroīdiem hormoniem gonadotropīna receptori ir saistīti ar mērķa šūnu membrānu. Peptīdu glikoproteīna hormonu virsmas šūnu receptori ir olbaltumvielas, kas ir daļa no šūnas membrānas struktūras. Pēc saistīšanās ar gonadotropīnu membrānas receptori stimulē šķīstošo intracelulāro kurjeru veidošanos, kas savukārt nodrošina šūnu reakciju (Halvorson L.M., ChinW.W., 1999).
Papildus hipotalāma liberīniem FSH ražošanas regulatori saskaņā ar mūsdienu koncepcijām ir inhibīns un aktivīns, ko ražo olnīcu granulozes un luteālās šūnas, kā arī citotrofoblastu šūnas (Hopko Ireland et al, 1994).
1. nodaļa. Sieviešu reproduktīvās sistēmas anatomija un fizioloģija 19
Inhibīns sastāv no divām kalmes apakšvienībām. FSH ietekmē inhibīna sintēzi un atbrīvošanos, izmantojot atgriezeniskās saites principu. oc-apakšvienības kombinācija ar 3-apakšvienību izraisa FSH nomākumu, un abu (3-apakšvienību) kombinācija izraisa aktivīna veidošanos un tādējādi FSH stimulāciju.
FSH sintēzi un izdalīšanos ietekmē arī follistatīns, kas izolēts no folikulu šķidruma. Follistatīns ir glikoproteīns, kas, tāpat kā inhibīns, samazina FSH izdalīšanos gonadotropo hipofīzes šūnu kultūrā. Turklāt tam ir augsta afinitāte pret aktivīna saistīšanos un mazāka pret inhibīna saistīšanos. Ir konstatēts, ka follistatīns un aktivīns A ir folikulu autokrīnās-parakrīnās sistēmas sastāvdaļas un ir iesaistīti Graafian pūslīšu iekšējās membrānas šūnu dažādu funkciju regulēšanā (Grome N., O "Brien ML, 1996).
Ir 3 gonadotropīna sekrēcijas veidi: tonizējoša, cikliska un epizodiska vai pulsējoša (Halvorson L.M., Chin W.W., 1999).
Tonizējošo jeb bazālo gonadotropīnu sekrēciju regulē negatīva atgriezeniskā saite, bet ciklisko - ar pozitīvas atgriezeniskās saites mehānismu, kurā iesaistīti estrogēni.
Pulsējoša sekrēcija ir saistīta ar hipotalāmu aktivitāti un gonadoliberīnu izdalīšanos.
Folikula attīstība cikla pirmajā pusē ir saistīta ar tonizējošu FSH un LH sekrēciju. Palielinot estradiola sekrēciju, tiek kavēta FSH veidošanās. Folikula attīstība ir atkarīga no FSH receptoru skaita granulozes zonas šūnās, un šo receptoru sintēzi savukārt stimulē estrogēni.
Tādējādi FSH noved pie estrogēnu sintēzes noteiktā folikulā, kas, palielinot FSH receptoru skaitu, veicina tā uzkrāšanos (saistoties ar tā receptoriem), folikulu turpmāku nobriešanu un estradiola sekrēcijas palielināšanos. Citi folikuli šajā laikā tiek pakļauti atrēzijai. Estradiola koncentrācija asinīs sasniedz maksimumu pirmsovulācijas periodā, kas izraisa liela daudzuma GnRH izdalīšanos un sekojošu LH un FSH izdalīšanās maksimumu. LH un FSH palielināšanās pirms ovulācijas stimulē Graafia pūslīšu plīsumu un ovulāciju (Hurk Van Den R., 1994).
LH ir galvenais steroīdu sintēzes regulators olnīcās. LH receptori ir lokalizēti luteālās šūnās, un LH iedarbība ir saistīta ar adenilātciklāzes stimulāciju un intracelulāru cAMP līmeņa paaugstināšanos, kas tieši vai caur starpniekiem (proteīnkināzi utt.) aktivizē progesterona biosintēzē iesaistītos enzīmus. . LH ietekmē olnīcās palielinās hormonu sintēzei nepieciešamā holesterīna daudzums. Tajā pašā laikā palielinās citohroma P450 saimes enzīmu aktivitāte, kas holesterīna molekulā atdala sānu ķēdi. Ar ilgāku iedarbību LH stimulē citu enzīmu ekspresiju un sintēzi (3V-hidroksisteroīdu dehidrogenāzes,
20 Endokrīnā ginekoloģija
17a-hidroksilāze), kas piedalās progesterona un citu steroīdu sintēzē. Tādējādi dzeltenajā ķermenī LH ietekmē pastiprinās steroidoģenēzes procesi holesterīna pārvēršanās vietā par pregnanolonu (Yen S., 1999).
Gonadotropīna sekrēcijas regulēšanu nodrošina "īsas" un "īpaši īsas" atgriezeniskās saites ķēdes. Tādējādi LH un FSH līmeņa paaugstināšanās izraisa to sintēzes un atbrīvošanās kavēšanu, un paaugstināta GnRH koncentrācija hipotalāmā kavē tā sintēzi un izdalīšanos hipofīzes portāla sistēmā (Sopelak V.M., 1997).
GnRH izdalīšanos ietekmē arī kateholamīni: dopamīns, adrenalīns un norepinefrīns. Epinefrīns un norepinefrīns stimulē GnRH izdalīšanos, savukārt dopamīnam ir tāda pati iedarbība tikai dzīvniekiem, kuriem iepriekš injicēti steroīdu hormoni. Holecistokinīns, gastrīns, neirotenzīns, opioīdi un somatostatīns kavē GnRH izdalīšanos (Yen S., 1999).
Adrenokortikotropajam hormonam ir stimulējoša iedarbība uz virsnieru garozu. Palielinot olbaltumvielu sintēzi (no cAMP atkarīgā aktivācija), rodas virsnieru garozas hiperplāzija. AKTH uzlabo holesterīna sintēzi un pregnanolona veidošanās ātrumu no holesterīna. Lielākā mērā tā iedarbība izpaužas uz fascikulāro zonu, kas izraisa glikokortikoīdu veidošanās palielināšanos, mazākā mērā - uz glomerulārās un retikulārās zonas, tāpēc tai nav būtiskas ietekmes uz mineralokortikoīdu veidošanos. un dzimumhormoni.
AKTH iedarbība ārpus virsnieru dziedzeriem ir lipolīzes stimulēšana (mobilizē taukus no tauku krātuvēm un veicina tauku oksidēšanos), pastiprināta insulīna un somatotropīna sekrēcija, glikogēna uzkrāšanās muskuļu šūnās, hipoglikēmija, kas saistīta ar pastiprinātu insulīna sekrēciju, pastiprināta pigmentācija. sakarā ar melanoforu iedarbību uz pigmenta šūnām .
Augšanas hormons ir iesaistīts augšanas un fiziskās attīstības regulēšanā, stimulējot olbaltumvielu veidošanos organismā, RNS sintēzi un aminoskābju transportēšanu no asinīm šūnās.
Prolaktīna galvenā bioloģiskā loma ir piena dziedzeru augšana un laktācijas regulēšana. Tas tiek darīts, stimulējot olbaltumvielu – laktalbumīna, tauku un piena ogļhidrātu sintēzi. Prolaktīns regulē arī dzeltenā ķermeņa veidošanos un tā progesterona veidošanos, ietekmē ūdens-sāļu metabolismu organismā, saglabājot organismā ūdeni un nātriju, pastiprina aldosterona un vazopresīna iedarbību, kā arī palielina tauku veidošanos no ogļhidrātiem.
Aizmugurējās hipofīzes hormoni tiek ražoti hipotalāmā. Neirohipofīzē tie uzkrājas. Hipotalāma supraoptisko un paraventrikulāro kodolu šūnās tiek sintezēts oksitocīns un antidiurētiskais hormons. Sintezētie hormoni tiek transportēti ar aksonu transportu ar neirofizīna nesējproteīna palīdzību pa hipotalāma-hipofīzes traktu uz hipofīzes aizmugurējo daivu. Šeit hormoni tiek nogulsnēti un pēc tam nonāk asinīs.
1. nodaļa. Sieviešu reproduktīvās sistēmas anatomija un fizioloģija 21
Antidiurētiskajam hormonam (ADH) jeb vazopresīnam organismā ir divas galvenās funkcijas. Tās antidiurētiskais efekts ir stimulēt ūdens reabsorbciju distālajā nefronā. Šī darbība tiek veikta, pateicoties hormona mijiedarbībai ar specifiskiem receptoriem, kā rezultātā palielinās cauruļveida sienas caurlaidība, tā reabsorbcija un urīna koncentrācija. Šajā gadījumā ūdens reabsorbcijas palielināšanās notiek arī hialuronidāzes aktivācijas dēļ kanāliņu šūnās, kas izraisa pastiprinātu hialuronskābes depolimerizāciju, kā rezultātā palielinās cirkulējošā šķidruma tilpums.
Lielās devās (farmakoloģiskās) ADH sašaurina arteriolus, kā rezultātā paaugstinās asinsspiediens. Tāpēc to sauc arī par vazopresīnu. Pie tā fizioloģiskās koncentrācijas asinīs šī darbība nav nozīmīga. ADH izdalīšanās palielināšanās, kas rodas asins zuduma, sāpju šoka laikā, izraisa vazokonstrikciju, kam šajos gadījumos ir adaptīva vērtība.
ADH ražošanas palielināšanās notiek, samazinoties ārpusšūnu un intracelulārā šķidruma tilpumam, samazinoties asinsspiedienam, paaugstinoties asins osmotiskajam spiedienam un aktivizējoties renīna-angiotenzīna un simpātiskās nervu sistēmas darbībai.
Oksitocīns selektīvi iedarbojas uz dzemdes gludajiem muskuļiem, izraisot to kontrakciju dzemdību laikā. Šis process tiek veikts, saistoties ar īpašiem oksitocīna receptoriem, kas atrodas uz šūnu virsmas membrānas. Augstas estrogēnu koncentrācijas ietekmē krasi palielinās receptoru jutība pret oksitocīnu, kas izskaidro dzemdes kontraktilās aktivitātes palielināšanos pirms dzemdībām.
Oksitocīna līdzdalība laktācijas procesā ir palielināt piena dziedzeru mioepitēlija šūnu kontrakciju, kā rezultātā palielinās piena sekrēcija. Savukārt oksitocīna sekrēcijas palielināšanās notiek impulsu ietekmē no dzemdes kakla receptoriem, kā arī krūšu sprauslu mehānoreceptoriem zīdīšanas laikā.
Nākamais reproduktīvās sistēmas līmenis ir olnīcas, kurās notiek steroīdu un folikuloģenēze, reaģējot uz gonadotropīnu ciklisku sekrēciju un augšanas faktoru (FR) ietekmē.
Olnīcas ir pārī savienots sievietes reproduktīvās sistēmas orgāns un tajā pašā laikā endokrīnais dziedzeris. Olnīca sastāv no diviem slāņiem: kortikālās vielas, kas pārklāta ar proteīna membrānu, un medulla. Atsevišķi aplūko olnīcu kaula daļu, kurā nav stromas teka-luteālās šūnas, kas satur granulētas šūnas, kas ir atbildīgas par olnīcu androgēnu veidošanos.
Garozu veido dažādas brieduma pakāpes folikuli (no pirmatnējā līdz atretiskajam), kas atrodas saistaudu stromā.
Folikuloģenēzes process olnīcā notiek nepārtraukti, un to regulē gonadotropīni, mijiedarbojoties ar olnīcu receptoriem (Sopelak V.M., 1997).
22 Endokrīnā ginekoloģija
Tajā pašā laikā katrā olnīcā tiek atklāti vairāki desmiti folikulu, kas atrodas dažādās augšanas un nobriešanas stadijās. Kopējais folikulu skaits dzimšanas brīdī ir aptuveni 2 miljoni.To skaits līdz menstruālā cikla izveidošanai samazinās 8-10 reizes, nepārsniedzot 30-40 tūkstošus.Tikai aptuveni 10% folikulu iziet pilnu attīstības ciklu no plkst. pirms ovulācijas un pārvēršas dzeltenā ķermenī. Pārējie tiek pakļauti atrēzijai un apgrieztai attīstībai (Hurk Van Den R. et al., 1994).
Primārā folikula transformācijas laikā par nobriedušu, tiek pabeigta pirmā mejozes dalīšanās, kuras rezultātā tiek atbrīvots vienvirziena (polārais) ķermenis un veidojas oocīts. Caurspīdīgais apvalks sasniedz maksimālo attīstību, pārvēršoties par mirdzošu vainagu, kas pārklāts ar 1-2 nejauši gulošu folikulu šūnu slāņiem. Folikulā veidojas dobums, kas sasniedz maksimālo izmēru pirms ovulācijas. Folikulāro šūnu slānis stromas asinsvadu augšanas faktoru ietekmē pārvēršas divos slāņos: folikula iekšējā un ārējā tekā. Turpmāka folikulu šķidruma daudzuma palielināšanās noved pie folikulu dobuma pārplūdes un tā plīsuma - ovulācijas. Pēc ovulācijas olšūna, ko ieskauj mirdzošs vainags, no vēdera dobuma nonāk olvadu piltuvē un pēc tam tās lūmenā. Šeit tiek pabeigta otrā mejozes dalīšanās un veidojas nobriedusi olšūna, kas ir gatava apaugļošanai (Yen S., 1999).
Olnīcu cikls sastāv no divām fāzēm - folikulārās un luteālās, kuras atdala ovulācija un menstruācijas.
Folikulārajā fāzē hipofīzes izdalītā FSH ietekmē kopā ar dažādiem augšanas faktoriem tiek stimulēta viena vai vairāku pirmatnējo folikulu augšana un attīstība, kā arī granulozes šūnu diferenciācija un proliferācija. FSH arī pastiprina 17-(3-hidroksisteroīdu dehidrogenāzes un aromatāzes aktivitāti, kas ir nepieciešamas estradiola veidošanai granulozes šūnās, aktivizējot cAMP), un tādējādi stimulē primāro folikulu augšanu un attīstību, folikulu epitēlija šūnu estrogēnu ražošanu. Est-radiols savukārt paaugstina granulozes šūnu jutību pret FSH darbību.FSH receptori pieder pie membrānas receptoru grupas ar 7 transmembrānu fragmentiem.Līdztekus estrogēniem izdalās neliels daudzums progesterona.No daudzajiem sākuma folikulu augšana, tikai 1 sasniegs galīgo briedumu, retāk - 2-3 .
Gonadotropīnu preovulācijas izdalīšanās nosaka ovulācijas procesu. Folikula tilpums strauji palielinās paralēli folikulu sieniņas retināšanai, kas saistīta ar proteolītisko enzīmu un polimorfonukleāro leikocītu izdalītās hialuronidāzes aktivitātes palielināšanos.
Novērots 2-3 dienu laikā pirms ovulācijas, ievērojams estrogēna līmeņa pieaugums ir saistīts ar liela skaita nobriedušu folikulu nāvi ar folikulu šķidruma izdalīšanos. Augstas estrogēnu koncentrācijas ar negatīvas atgriezeniskās saites mehānismu kavē FSH sekrēciju no hipofīzes. Ovulācijas LH pieaugums un mazākā mērā
1. nodaļa. Sieviešu reproduktīvās sistēmas anatomija un fizioloģija 23
FSH pakāpe ir saistīta ar pozitīvas atgriezeniskās saites mehānisma esamību īpaši augstas estrogēnu un LH koncentrācijas gadījumā, kā arī ar strauju estradiola līmeņa pazemināšanos 24 stundu laikā pirms ovulācijas.
Menstruālā cikla neirohormonālā regulēšana shematiski parādīta 6. attēlā.
I Ovulācija

6. attēls. Menstruālā cikla neirohormonālā regulēšana
Olas ovulācija notiek tikai LH vai cilvēka horiona gonadotropīna klātbūtnē. Turklāt FSH un LH darbojas kā sinerģisti folikulu attīstības laikā, kura laikā teka šūnas aktīvi izdala estrogēnus.
Folikula sienas kolagēna slāņa iznīcināšanas mehānisms ir no hormoniem atkarīgs process, kura pamatā ir folikulu fāzes atbilstība. LH preovulācijas pieaugums stimulē progesterona koncentrācijas palielināšanos ovulācijas laikā. Sakarā ar pirmo progesterona maksimumu palielinās folikulu sieniņas elastība, līdz ar to FSH, LH un progesterons kopīgi stimulē proteolītisko enzīmu darbību: granulozes šūnu izdalītie plazminogēna aktivatori veicina plazmīna veidošanos, plazmīns ražo dažādas kolagenāzes, prostaglandīnus E un F2ot veicina šūnu oocītu masas uzkrāšanās pārvietošanu. Lai novērstu neovulējošā folikula priekšlaicīgu luteinizāciju, olnīcā ir jāveido noteikts aktivīna daudzums (Speroff L. et al., 1994).
Pēc ovulācijas asins serumā strauji pazeminās LH un FSH līmenis. No cikla otrās fāzes 12.dienas par 2-3 dienām paaugstinās FSH līmenis asinīs, kas ierosina jauna folikula nobriešanu, savukārt LH koncentrācijai ir tendence samazināties visā otrajā. cikla fāze.
Kovulētā folikula dobums sabrūk, un tā sienas sakrājas krokās. Asinsvadu plīsuma dēļ ovulācijas laikā pēcovulācijas folikula dobumā rodas asiņošana. Topošā dzeltenā ķermeņa centrā parādās saistaudu rēta - stigma (Speroff L. et al., 1994).
24 Endokrīnā ginekoloģija
LH ovulācijas pieaugums un sekojoša augsta hormona līmeņa uzturēšana 5-7 dienas aktivizē granulētās zonas šūnu (granulozes) proliferācijas un dziedzeru metamorfozes procesu, veidojot luteālās šūnas, t.i. sākas olnīcu cikla luteālā fāze (dzeltenā ķermeņa fāze) (Erickson G.F., 2000).
Folikula granulētā slāņa epitēlija šūnas intensīvi vairojas un, uzkrājot lipohromus, pārvēršas par luteālās šūnām; pati čaula ir bagātīgi vaskularizēta. Vaskularizācijas stadijai raksturīga strauja granulozes epitēlija šūnu pavairošana un intensīva kapilāru augšana starp tām. Asinsvadi iekļūst pēcovulācijas folikula dobumā no thecae internae puses luteālās audos radiālā virzienā. Katra dzeltenā ķermeņa šūna ir bagātīgi apgādāta ar kapilāriem. Saistaudi un asinsvadi, sasniedzot centrālo dobumu, piepilda to ar asinīm, apņem pēdējo, ierobežojot to no luteālo šūnu slāņa. Dzeltenajā ķermenī ir viens no augstākajiem asinsrites līmeņiem cilvēka organismā. Šī unikālā asinsvadu tīkla veidošanās beidzas 3-4 dienu laikā pēc ovulācijas un sakrīt ar dzeltenā ķermeņa funkcijas ziedu laikiem (Bagavandoss P., 1991).
Angioģenēze sastāv no trim fāzēm: esošās bazālās membrānas fragmentācijas, endotēlija šūnu migrācijas un to proliferācijas, reaģējot uz mitogēnu stimulu. Angiogēno aktivitāti kontrolē galvenie augšanas faktori: fibroblastu augšanas faktors (FGF), epidermas augšanas faktors (EGF), trombocītu augšanas faktors (PGF), insulīnam līdzīgs augšanas faktors-1 (IGF-1), kā arī citokīni, piemēram, kā nekrotiskā faktora audzējs (TNF) un interleikīni (IL-1; IL-6) (Bagavandoss P., 1991).
No šī brīža dzeltenais ķermenis sāk ražot ievērojamu daudzumu progesterona. Progesterons īslaicīgi inaktivē pozitīvās atgriezeniskās saites mehānismu, un gonadotropīnu sekrēciju kontrolē tikai zstradiola negatīvā iedarbība. Tas noved pie gonadotropīnu līmeņa pazemināšanās dzeltenā ķermeņa fāzes vidū līdz minimālajām vērtībām (Erickson G.F., 2000).
Progesterons, ko sintezē dzeltenā ķermeņa šūnas, kavē jaunu folikulu augšanu un attīstību, kā arī piedalās endometrija sagatavošanā apaugļotas olšūnas ievadīšanai, samazina miometrija uzbudināmību, nomāc estrogēnu ietekmi uz endometrijs cikla sekrēcijas fāzē, stimulē deciduālo audu attīstību un alveolu augšanu piena dziedzeros. Progesterona koncentrācijas līmenis serumā atbilst rektālās (bazālās) temperatūras plato (37,2-37,5 ° C), kas ir vienas no notikušās ovulācijas diagnostikas metodēm un ir kritērijs luteālās zarnas lietderības novērtēšanai. fāze. Bāzes temperatūras paaugstināšanās pamatā ir perifērās asins plūsmas samazināšanās progesterona ietekmē, kas samazina siltuma zudumus. Tā satura palielināšanās asinīs sakrīt ar bazālās temperatūras paaugstināšanos, kas ir ovulācijas rādītājs (McDonnel D.P., 2000).
1. nodaļa. Sieviešu reproduktīvās sistēmas anatomija un fizioloģija 25
Progesterons, būdams estrogēnu antagonists, ierobežo to proliferatīvo efektu endometrijā, miometrijā un maksts epitēlijā, izraisot glikogēnu saturošas sekrēcijas stimulāciju endometrija dziedzeros, samazinot submukozālā slāņa stromu, t.i. izraisa raksturīgas izmaiņas endometrijā, kas nepieciešamas apaugļotas olšūnas implantācijai. Progesterons samazina dzemdes muskuļu tonusu, liekot tiem atpūsties. Turklāt progesterons izraisa piena dziedzeru proliferāciju un attīstību un grūtniecības laikā veicina ovulācijas procesa kavēšanu (O "Malleu B.W., Strott G.A., 1999).
Šīs folikulu attīstības fāzes ilgums ir atšķirīgs: ja apaugļošanās nenotiek, tad pēc 10-12 dienām menstruālais dzeltenais ķermenis regresē, ja apaugļotā olšūna ir iekļuvusi endometrijā un iegūtā blastula sāk sintezēt horiona gonadotropīnu (CG), tad dzeltenais ķermenis kļūst par grūtniecības dzelteno ķermeni.
Dzeltenā ķermeņa granulozes šūnas izdala polipeptīdu hormonu relaksīnu, kam ir svarīga loma dzemdību laikā, izraisot iegurņa saišu relaksāciju un dzemdes kakla relaksāciju, kā arī palielina glikogēna sintēzi un ūdens aizturi miometrijā, vienlaikus samazinot tā kontraktilitāti. Parastā menstruālā cikla laikā tā sekrēcija palielinās uzreiz pēc LH izdalīšanās maksimuma un paliek nosakāma menstruāciju laikā. Grūtniecības laikā cirkulējošais relaksīna līmenis pirmā trimestra beigās ir augstāks nekā otrajā un trešajā trimestrī.
Ja olšūna apaugļošanās nenotiek, dzeltenais ķermenis nonāk apgrieztās attīstības stadijā, ko pavada menstruācijas. Luteālās šūnās notiek distrofiskas izmaiņas, samazinās izmērs, savukārt tiek novērota kodolu piknoze. Saistaudi, augot starp trūdošajām luteālās šūnām, tos nomaina, un dzeltenais ķermenis pamazām pārvēršas par hialīna veidojumu - balto ķermeni (corpus albicans) (Sopelak V.M., 1997).
No hormonālās regulēšanas viedokļa dzeltenā ķermeņa regresijas periodam ir raksturīgs izteikts progesterona, estradiola un inhibīna A līmeņa pazemināšanās. Inhibīna A līmeņa pazemināšanās novērš tā bloķējošo ietekmi uz hipofīzi. dziedzeri un FSH sekrēcija. Tajā pašā laikā progresējoša estradiola un progesterona koncentrācijas samazināšanās veicina strauju GnRH sekrēcijas biežuma palielināšanos, un hipofīze tiek atbrīvota no negatīvās atgriezeniskās saites kavēšanas. Inhibīna A un estradiola līmeņa pazemināšanās, kā arī Gn-RH sekrēcijas impulsu biežuma palielināšanās nodrošina FSH sekrēcijas pārsvaru pār LH. Reaģējot uz FSH līmeņa paaugstināšanos, beidzot veidojas antrālo folikulu kopums, no kura turpmāk tiks izvēlēts dominējošais folikuls. Prostaglandīnam F2a, oksitocīnam, citokīniem, prolaktīnam un 02 radikāļiem ir luteolītiska iedarbība, kas var būt par pamatu dzeltenā ķermeņa nepietiekamības attīstībai piedēkļu iekaisuma procesa klātbūtnē.
Olnīcu (menstruālā) cikla ilgums parasti svārstās no 21 līdz 35 dienām.
Menstruācijas notiek uz dzeltenā ķermeņa regresijas fona. Līdz tā beigām estrogēna un progesterona līmenis sasniedz minimālo līmeni. Uz šī fona, tur
26 Endokrīnā ginekoloģija
Hipotalāma un hipofīzes tonizējošā centra aktivizēšana un pastiprināta galvenokārt FSH sekrēcija, kas aktivizē folikulu augšanu. Estradiola līmeņa paaugstināšanās izraisa proliferācijas procesu stimulāciju endometrija bazālajā slānī, kas nodrošina adekvātu endometrija atjaunošanos (7. att.).

7. attēls. Normāla menstruālā cikla regulēšanas saites (Sopelak V., 1997)
1. nodaļa. Sieviešu reproduktīvās sistēmas anatomija un fizioloģija 27
Olnīcu steroidoģenēze notiek epitēlija šūnās, kas pārklāj folikulu dobumu, iekšējās tekas šūnās un daudz mazāk stromā. Folikulārās epitēlija šūnas, stromas un teka audi sintezē progesteronu, testosteronu, dehidrotestosteronu, estronu un estradiolu (Erickson G.F., 2000).
Estrogēni ir estradiols, estrons un estriols. Bioloģiski aktīvākais ir estradiols, no kura 95% veidojas folikulā, un tā līmenis asinīs ir folikula nobriešanas indikators. Estradiolu (E2) galvenokārt izdala granulozes šūnas un mazākā mērā dzeltenais ķermenis. Estronu (E,) veido estradiola perifēra aromatizēšana. Galvenais estriola (E3) avots ir estradiola un estrona hidroksilēšana aknās (O "Malleu B.W., Strott G.A., 1999).
Asinīs izdalītos estrogēnus konjugē ar sekssteroīdu saistošo globulīnu (SHBG) un mazākā mērā ar asins albumīnu. SHBG ir pazīstams arī kā estradiolu-testosteronu saistošs globulīns. Pats nosaukums norāda uz šī proteīna palielināto afinitāti pret androgēniem. Dzimumhormonus saistošā globulīna līmenis sieviešu asins serumā ir gandrīz 2 reizes augstāks, salīdzinot ar tā koncentrāciju vīriešu asinīs. Estrogēni un to metabolīti tiek konjugēti aknās ar glikuronskābi un sērskābi un izdalās ar žulti un urīnu (McDonnel D.P., 2000).
Papildus jau minētajai ietekmei uz dzimumorgāniem, hipofīzi un hipotalāmu, estrogēniem piemīt anaboliska īpašība, tie palielina kaulu audu vielmaiņu un paātrina skeleta kaulu nobriešanu, kas ir iemesls augšanas pārtraukšanai plkst. pubertātes sākums, no vienas puses, un juvenīlās osteoporozes attīstība meitenēm ar aizkavētu seksuālo attīstību – no otras puses.
Lielās devās estrogēni veicina nātrija un ūdens aizturi organismā līdz tūskas attīstībai. Tie ietekmē arī lipīdu metabolismu, pazeminot holesterīna līmeni asinīs.
Progesteronu izdala dzeltenais ķermenis, kā arī virsnieru garoza un sēklinieki, kur to izmanto kā kortikosteroīdu un androgēnu biosintēzes prekursoru. Progestogēniem un glikokortikoīdiem ir līdzīga ķīmiskā struktūra, tāpēc progesterona un glikokortikoīdu receptoriem ir savstarpējas saites īpašības. Serumā progesteronu saistās ar transkortīnu, kas, kā zināms, saistās arī ar glikokortikoīdiem. Saskaņā ar dažiem pētījumiem progesterona spēja saistīt transkortīnu pat pārsniedz kortikosteroīdu spēju. Aknās progesterons saistās ar glikuronskābi un konjugētā veidā izdalās ar urīnu (McDonnel D.P., 2000). Taču estrogēnu un progesterona ietekme uz mērķorgāniem sīkāk aprakstīta sadaļā "Dzimuma steroīdu hormonu lietošanas principi klīniskajā praksē un to sistēmiskā iedarbība".
Androgēnus sievietēm izdala olnīcu stromas šūnas, galvenokārt androstenediona veidā, un virsnieru dziedzeros tas veidojas 3 reizes vairāk nekā olnīcās. Androstenedions perifēros audos tiek pārveidots par testosteronu. Olnīcās tas veidojas mazos
28 Endokrīnā ginekoloģija
Daudzumos arī testosterons, dihidrotestosterons, dehidroepiandrosterons. Apmēram 1/4 testosterona, kas izdalās sievietes ķermenī, tiek ražots olnīcās. Pārējo tā daudzumu izdala virsnieru dziedzeri vai veidojas audos perifērijā, pārvēršoties no androstenediona (McDonnel D.P., 2000).
Steroīdu bioloģiskā iedarbība mērķa audos ir saistīta ar specifisku receptoru klātbūtni tajos (8. att.). Steroīdi izkliedējas caur šūnu membrānu un saistās ar specifiskiem receptoriem citoplazmā. Steroīdu receptori ir salīdzinoši lieli proteīni ar augstu saistīšanās spēju ar noteiktiem hormoniem. Tomēr ir iespējama šo receptoru saistīšanās ar citiem šīs grupas steroīdiem (piemēram, sintētiskiem agonistiem un antagonistiem). Citoplazmas receptori nav visās, bet tikai audu šūnās, kas ir jutīgas pret šāda veida hormoniem. Steroīdu-receptoru komplekss, kura veidošanās ir atkarīga no vairākiem faktoriem, tostarp temperatūras, pārvietojas uz kodolu, kur uz hromatīna ir īpašas vietas, kas saista šos kompleksus. Steroīdu-receptoru komplekss tiek aktivizēts, pēc tam tas var saistīties ar akceptora kodolproteīnu, kas atrodas uz DNS. Pēdējā mijiedarbība noved pie liela skaita specifisku RNS un atbilstošu proteīnu sintēzes, atbilstošo orgānu (piena dziedzeru, dzemdes utt.) un audu augšanas un attīstības (O "Malleu B.W., Strott G.A., 1999).
![]()
8. attēls. Steroīdu hormonu darbības mehānisms uz mērķa audiem (Cowan B.D., 1997)
Dažādu steroīdu hormonu receptoru molekulu skaits svārstās no 5000 līdz 20 000 vienā šūnā. Estrogēnu receptori saista daudzus
1. nodaļa. Sievietes reproduktīvās sistēmas anatomija un fizioloģija 29
Gee dabiskie un sintētiskie estrogēnie steroīdi ar tādu pašu afinitāti. Tiek uzskatīts, ka estrogēna un progesterona receptori ir divas apakšvienības, no kurām katra saista hormona molekulu, kā tas ir sīkāk aprakstīts klīniskajā nodaļā "Dzimuma steroīdu hormonu lietošanas principi klīniskajā praksē".
Katra no a un P apakšvienībām mijiedarbojas ar hromatīnu un nodrošina specifisku gēnu un RNS polimerāžu turpmāku aktivāciju.
Hormona bioloģiskā iedarbība ir saistīta ne tikai ar tā kvantitatīvajām svārstībām asins serumā, bet arī ar receptoru saites stāvokli, un receptoru skaits ir pakļauts ievērojamām svārstībām. Eksperimentālie pētījumi ir parādījuši, ka jaundzimušo žurku mērķa audi satur nelielu daudzumu estrogēnu receptoru. 10. dzīves dienā receptoru skaits palielinās, un pēc šī perioda eksogēno estrogēnu ievadīšana izraisa to palielināšanos. Estrogēni stimulē ne tikai estrogēna, bet arī progesterona receptoru veidošanos. Receptoru skaits ir atkarīgs ne tikai no hormona līmeņa, kas cirkulē asinīs, bet ir arī ģenētiskā kontrolē. Tādējādi sēklinieku feminizācijas sindromā tiek novērota androgēnu receptoru pilnīga neesamība (McDonnel D.P., 1999).

9. attēls. Steroīdu hormonu ķīmiskā struktūra (Sopelak V., 1997)
Galveno dzimumhormonu ķīmiskās struktūras analīze liecina, ka tie visi ir progesterona atvasinājumi, un estrogēni cits no cita atšķiras tikai ar to struktūrā esošo hidroksiradikāļu skaitu (9. att.).
30 Endokrīnā ginekoloģija
Visu steroīdu hormonu viela ir zema blīvuma lipoproteīnu holesterīns (ZBL). Steroidoģenēzē ir iesaistīti gonadotropīni (FSH un LH), kā arī enzīmu sistēmas (aromātāzes). Pirmkārt, pregnanolons veidojas holesterīna sānu ķēdes šķelšanās rezultātā. Nākotnē ir iespējami divi pregnanolona metabolisko transformāciju veidi, kas beidzas ar testosterona veidošanos, kas saņēma m- un n5-metabolisma ceļu nosaukumus, pamatojoties uz dubultās nepiesātinātās saites stāvokli iegūtajos savienojumos. Dominējošā dzimuma steroīdu veidošanās notiek pa L5 ceļu. Tās gaitā secīgi veidojas 17a-hidroksipregnanolons, dehidroepiandrosterons (DHEA), androstenedions. Progesterons, 17a-hidroksiprogesterons un androstenedions veidojas pa L4 ceļu. A4,5-izomerāze aizver abus ceļus. Tālāk notiek testosterona vai androstenediona aromatizācija, veidojot attiecīgi estradiolu vai estronu (10. att.).

Piezīme: GSD - 3p-hidroksisteroīdu dehidrogenāze, DOC - deoksikortikosterons
10. attēls. Steroīdu biosintēze (Cowan B.D., 1997)
Lielākā daļa steroidogēno enzīmu, kas pārvērš holesterīnu prekursoros un bioloģiski aktīvos steroīdos, pieder P450 citohromu grupai. Citohroms P450 ir vispārējs termins daudziem oksidatīviem enzīmiem (Bryan D., 1997). Ir aptuveni 200 citohromu veidu, no kuriem pieci ir iesaistīti steroidoģenēzes procesā (2. tabula).
Procesā iesaistītie P450 fermenti 2. tabula
Steroidoģenēze
![]()
1. nodaļa. Sieviešu reproduktīvās sistēmas anatomija un fizioloģija 31
Reproduktīvās sistēmas perifēro saiti pārstāv mērķa orgāni, kas ietver dzimumorgānus un piena dziedzerus, kā arī ādu un tās piedēkļus, kaulus, asinsvadus un taukaudi. Šo audu un orgānu šūnās atrodas dzimumhormonu receptori, kas ir citoplazmas receptori – citozola receptori. Tāpat dzimumhormonu receptori ir atrodami visās reproduktīvās sistēmas struktūrās un, pats galvenais, centrālajā nervu sistēmā (McDonnel D.P., 2000).
Tādējādi reproduktīvā sistēma ir vienota neatņemama sistēma, kuras visas saites ir savstarpēji saistītas gan ar tiešās, gan atgriezeniskās saites mehānismu.
Literatūra
1. Bagavandoss P, Vilkss, Dž. Mikrovaskulāro endotēlija šūnu izolēšana un raksturojums no jaunattīstības dzeltenā ķermeņa. Biol. Reprod 1991; 44: 1132-1139.
2. Braiens D. Steroīdu biosintēze / Bryan D. Cowan, David B. Seifer Klīniskā reproduktīvā medicīna. Filadelfija-Ņujorka 1997: 11-20.
3. Cowan B.D. Steroīdu biosintēze. Klīniskā reproduktīvā medicīna / Ed Cowan BD, Seifer DB. Filadelfija-Ņujorka: Lippincott-Raven Publishers 1997: 11-20.
4. Ēriksons G.F. Olnīcu anatomija un fizioloģija. menopauze. Bioloģija un patobioloģija / Ed Lobo RA, Kelsey J, Marcus R. San Diego: Academic Press 2000: 13-32.
5. Gougeon A. Olnīcu folikulu attīstības regulēšana primātiem: fakti un hipotēze. Endokr. Rev 1996; 17:121-155.
6. Grome N, O "Braiens M. Dimēra inhibīcijas B mērīšana menstruālā cikla laikā. J. Clin. Endocr. Metab, 1996; 81: 1400-1405.
7. Halvorson LM, Chin WW. Gonadotropiskie hormoni: byosintēze, sekrēcija, receptori un darbība. Reproduktīvā endokrinoloģija / Ed Yen SSC, Jaffe RB, Barbieri RL, Filadelfija, ASV 1999: 30-80.
8. Hopko Ireland, Janet L, Ireland JJ. Izmaiņas inhibīna/aktivīna un subvienību vēstnesis ribonukleīnskābju ekspresijā pēc izmēra palielināšanās un dažādos neovulācijas folikulu diferenciācijas vai atrēzijas posmos govīm. Biol Reprod 1994; 50:492-501.
9. Hurk Van Den R, Dijkstra G, Hulshof SCJ, Vos PLAM. Liellopu antrālo folikulu mikromorfoloģija pēc prostaglandīnu izraisītas luteolīzes, īpaši atsaucoties uz netipiskām granulozes šūnām. J Reprod Fertil 1994; 100:137-142.
10. Lakoskis JM. Šūnu elektrofizioloģiskās pieejas sieviešu reproduktīvās novecošanas centrālajai regulēšanai. Reproduktīvās funkcijas neironu kontrole / Ed Lakoski JM, Perez-Polo JR, Rassin DK. Ņujorka: Liss 1989: 209-220.
11. McDonnel D.P. Estrogēna un progesterona receptoru molekulārā farmakoloģija. menopauze. Bioloģija un patobioloģija / Ed Lobo RA, Kelsey J, Marcus R. San Diego: Academic Press 2000: 3-12.
12. O "Malleu BW, Strott GA. Steroīdu hormoni: vielmaiņa un darbības mehānisms. Reproduktīvā endokrinoloģija / Ed Yen SSC, Jaffe RB, Barbieri RL, Filadelfija, ASV 1999: 110-133.
13. Sopelak VM. HPO ass neiroregulācija / Ed Bryan D. Cowan, David B. Seifer. Klīniskā reproduktīvā medicīna. Filadelfija-Ņujorka 1997: 3-10.
14. Sopelak VM. Olnīcu-menstruālā cikla regulēšana / Ed Bryan D. Cowan, David B. Seifer. Klīniskā reproduktīvā medicīna. Filadelfija-Ņujorka 1997: 61-68.
32 Endokrīnā ginekoloģija
15. Speroff L, Glass NG, Kase. Klīniskā ginekoloģiskā endokrinoloģija un neauglība, 1994: 213-220.
16. Wildt L. Hipotalāms. Reprodukcijasmedizin / Ed Hrsg. von Bettendorf G, Breckwoldt M. Stuttgart: Fischer 1989: 6-22.
17. Yen S.S.C. Cilvēka menstruālais cikls: neiroendokrīnā regulēšana. Reproduktīvā endokrinoloģija / Ed Yen SSC, Jaffe RB, Barbieri RL. Filadelfija, ASV 1999: 191-217.
Jenas SSC. Reprodukcijas neiroendokrinoloģija. Reproduktīvā endokrinoloģija / Ed Yen SSC, Jaffe RB, Barbieri RL. Filadelfija, ASV 1999: 30-80.
Saistītie raksti