硬骨魚の一般的な特徴。 骨。 骨組織の機能。 骨組織の構造 骨組織の特徴
記事の内容
骨、脊椎動物にのみ特徴的な高密度の結合組織。 骨は身体を構造的に支え、身体全体の形と大きさを維持するのに役立ちます。 一部の骨の位置は、脳などの軟部組織や臓器を保護し、獲物の硬い殻を破ることができない捕食者による攻撃に抵抗するような位置にあります。 骨は四肢に強度と剛性を与え、筋肉の付着部位としても機能し、四肢が移動や採餌という重要な機能においてレバーとして機能できるようにします。 最後に、骨には鉱床が多く含まれているため、無機物質が蓄えられ、必要に応じて消費されます。 この機能は、血液や他の組織のカルシウムバランスを維持するために非常に重要です。 あらゆる臓器や組織でカルシウムの必要性が突然増加すると、骨がカルシウムの補給源となることがあります。 したがって、一部の鳥では、卵の殻の形成に必要なカルシウムが骨格から得られます。
骨格系の古さ。
この骨は、知られている最古の化石脊椎動物であるオルドビス紀(約5億年前)の装甲を備えた無顎動物の骨格に存在している。 これらの魚のような生き物では、骨は体を保護する外側の板の列を形成する役割を果たしました。 それらのいくつかは、頭部の内部骨骨格も持っていましたが、内部骨骨格の他の要素は持っていませんでした。 現代の脊椎動物の中には、骨が完全に、またはほぼ完全に存在しないことを特徴とするグループがあります。 しかし、それらのほとんどについては、過去に骨骨格が存在したことが知られており、現代の形態に骨が存在しないのは、進化の過程で骨が減少(喪失)した結果です。 たとえば、現生のサメのすべての種には骨がなく、軟骨に置き換えられています(鱗の基部と背骨には非常に少量の骨組織があり、主に軟骨で構成されています)が、その祖先の多くは現在、絶滅しており、発達した骨骨格を持っていました。
骨の本来の機能はまだ正確に解明されていません。 古代の脊椎動物のそれらのほとんどが体の表面またはその近くに位置していたという事実から判断すると、この機能が補助的であったとは考えられません。 一部の研究者は、骨の本来の機能は、鎧を着た最古の無顎類を甲殻類(ユーリプテリド類)などの大型無脊椎動物の捕食者から守ることであったと考えています。 言い換えれば、外骨格は文字通りの鎧の役割を果たしました。 すべての研究者がこの観点を共有しているわけではありません。 古代の脊椎動物における骨のもう一つの機能は、現代の多くの脊椎動物で観察されているように、体内のカルシウムバランスを維持することであった可能性があります。
細胞間骨物質。
ほとんどの骨は、細胞によって生成される高密度の細胞間骨物質の中に点在する骨細胞 (骨細胞) で構成されています。 細胞は骨の総体積のほんの一部しか占めず、一部の成体脊椎動物、特に魚類では細胞間物質の生成に寄与した後に細胞が死亡するため、成熟した骨には細胞が存在しません。
骨の細胞間空間は、有機物質とミネラルという 2 つの主要な種類の物質で満たされています。 細胞活動の結果である有機塊は、主にタンパク質(束を形成するコラーゲン線維を含む)、炭水化物、および脂質(脂肪)で構成されます。 通常、骨物質の有機成分の大部分はコラーゲンです。 一部の動物では、骨物質の体積の 90% 以上を占めます。 無機成分は主にリン酸カルシウムに代表される。 正常な骨形成中、カルシウムとリン酸塩は血液から発育中の骨組織に入り、骨細胞によって生成される有機成分とともに骨の表面と深さに沈着します。
成長と老化に伴う骨組成の変化に関する私たちの知識のほとんどは、哺乳類の研究から得られています。 これらの脊椎動物では、有機成分の絶対量は生涯を通じて多かれ少なかれ一定ですが、ミネラル(無機)成分は年齢とともに徐々に増加し、成体では骨格全体の乾燥重量のほぼ65%を占めます。 。
物理的特性
骨は体を保護し、支える機能に適しています。 骨は強くて硬いと同時に、通常の生活条件下で折れないほど十分な弾力性がなければなりません。 これらの特性は、細胞間骨物質によって提供されます。 骨細胞自体の寄与は重要ではありません。 剛性、つまり 曲げ、伸張、または圧縮に抵抗する能力は、有機成分、主にコラーゲンによって提供されます。 後者は骨に弾力性を与えます。これは、わずかな変形(曲げまたはねじれ)の場合に元の形状と長さを復元できる特性です。 細胞間物質の無機成分であるリン酸カルシウムも骨の硬さに寄与しますが、主に骨に硬さを与えます。 特別な処理によってリン酸カルシウムが骨から除去されると、その形状は保たれますが、硬度は大幅に失われます。 硬度は骨の重要な性質ですが、残念なことに、硬度が過度の負荷がかかると骨を骨折しやすくします。
骨の分類。
骨の構造は、生物が異なれば、また同じ生物の体の部位によっても大きく異なります。 骨は密度によって分類できます。 骨格の多くの部分 (特に長骨の骨端)、特に胎児骨格では、骨組織には、緩い結合組織や血管で満たされた多くの空隙や溝があり、横木や支柱のネットワークとして現れます。金属橋の構造。 このような骨組織によって形成された骨は海綿状と呼ばれます。 身体が成長するにつれて、疎性結合組織と血管によって占められているスペースの多くが追加の骨物質で満たされ、その結果骨密度が増加します。 狭い管が比較的まばらなこの種の骨は、緻密または緻密と呼ばれます。
成体生物の骨は、周囲に沿って位置する緻密で緻密な物質と、中心に位置する海綿状物質で構成されています。 異なる種類の骨におけるこれらの層の比率は異なります。 したがって、海綿骨では、緻密層の厚さは非常に薄く、主な質量は海綿状物質によって占められています。
骨は、細胞間物質内の骨細胞の相対的な数と位置、およびこの物質の重要な部分を構成するコラーゲン束の方向によっても分類できます。 で 管状の骨では、コラーゲン線維の束がさまざまな方向に交差しており、骨細胞は細胞間物質全体に多かれ少なかれランダムに分布しています。 フラット骨はより秩序だった空間構成を持ち、連続した層 (プレート) で構成されています。 単一層の異なる部分では、コラーゲン線維は通常同じ方向を向いていますが、隣接する層では異なる場合があります。 扁平骨は管状骨よりも骨細胞が少なく、層内と層間の両方に存在します。 骨性骨は平らな骨と同様に層状構造をしていますが、その層はいわゆる狭い周りの同心円状のリングです。 血管が通るハバース管。 層は外側の層から形成され、そのリングは徐々に狭くなり、チャネルの直径が減少します。 ハバース運河と周囲の層はハバースシステムまたはオステオンと呼ばれます。 骨形成性骨は、通常、海綿骨から緻密骨への移行中に形成されます。
表面膜と骨髄。
密集した骨が関節で接触し、軟骨で覆われている場合を除き、骨の外面と内面は緻密な膜で覆われており、これは骨の機能と安全性に不可欠です。 外膜は骨膜または骨膜(ギリシャ語から)と呼ばれます。 ペリ- その周り、 オステオン- 骨)、骨空洞に面した内部のものは、内部骨膜または骨内膜(ギリシャ語から。 オンドン- 内部)。 骨膜は 2 つの層で構成されています。外側の線維 (結合組織) 層は、弾性のある保護殻であるだけでなく、靱帯や腱の付着部位でもあります。 そして骨の厚みを確実に成長させる内層。 骨内膜は骨の修復にとって重要であり、骨膜の内層にある程度似ています。 成長と骨吸収の両方をもたらす細胞が含まれています。
多くの骨、特に四肢の骨、椎骨、肋骨、骨盤の深部には、体の主な血球供給源である骨髄があります。 哺乳類を含む多くの脊椎動物の胎生期および出生直後には、骨髄 (赤色) がほぼすべての骨に存在し、造血細胞が非常に豊富です。 年齢とともに骨髄の造血活動は低下し、脂肪細胞(黄色の骨髄)が主成分になります。
細胞要素と骨の発達。
動物の一生を通して、骨は常に新しくなります。 多くの骨、特に発生初期に形成される骨は、あらゆる種類の結合組織の源である未分化の間葉細胞から形成されます。 将来の骨の局在化の場所では、間葉細胞のグループが徐々に分化し、細胞間骨物質の有機成分を活発に生産および分泌し始めます。 これらの細胞は骨芽細胞と呼ばれます。 有機成分が形成された後、石灰化が始まり、リン酸カルシウムが沈着します。 後の段階では、骨芽細胞は成熟した骨細胞、つまり骨細胞に変化します。 骨細胞の主な機能は、組織の石灰化を望ましいレベルに維持することです。 このようにして、いわゆる現像が起こる。 頭頂骨や前頭骨などの主要な骨。 子宮内発育の後期段階で起こる管状骨やその他の(二次)骨の形成は、異なる方法で進行します。まず、将来の骨の成長中の軟骨モデルが形成され、次に胎児の発育に伴って、また胎児の成長後にも形成されます。子供の誕生により、軟骨は徐々に骨組織に置き換わります。 骨組織の吸収は、血液単球から発生する特別な種類の骨マクロファージである破骨細胞によって確実に行われます。 破骨細胞は、骨物質を効果的に溶解して破壊する酵素を生成します。
骨の改造。
ほとんどすべての骨は、動物の成長中にその形状を変化させます。これは、ある場所で骨が成長し、別の場所で骨が破壊されることによって達成されます。 たとえば、手足の骨は長さだけでなく幅も大きくなります。 骨膜は骨芽細胞の供給源であり、骨芽細胞は外表面への骨組織の堆積を保証し、一方、骨内膜破骨細胞は骨を破壊して吸収し、それによって髄腔を拡張します。 全体的な成長がない場合でも、骨組織の継続的な再構成が発生します。古い骨組織は吸収され、新しい骨組織に置き換わります。 たとえば、犬では、骨組織の最大 10% が毎年入れ替わります。
骨のリモデリングは、体重により圧力が上昇する領域での骨の成長などの機能変化に応じて定期的に発生します。 また、損傷後の骨の修復、特に骨折の際、創傷の一次治癒に続いて再構築が行われ、徐々に骨の元の形状を回復する際にも主導的な役割を果たします。
血液供給
骨の形成に重要です。 間葉細胞の骨芽細胞への分化は、毛細血管血流の存在下でのみ起こります。 毛細血管を失った間葉は、軟骨組織を生成する細胞に変わります。 骨(特に骨)は血管の周囲に堆積することが多いため、多くの骨格骨の三次元組織構造の形成を決定します。
病気。
骨疾患は、骨の成長と再構築に伴う 3 つの主要なプロセスすべてを混乱させる可能性があります。 骨基部の石灰化。 破骨細胞による骨吸収。 壊血病は、骨組織の有機成分であるコラーゲンの生成を阻害することによる骨の成長への影響など、さまざまな結合組織に影響を与えます。 石灰化は直接影響を受けないため、生成される少量の有機物の過剰な石灰が発生します。 骨の成長はほぼ完全に止まり、非常にもろくなってしまいます。 逆に、くる病(子供が罹患する)や骨軟化症(成人の病気)では、石灰化が著しく損なわれます。 骨芽細胞はコラーゲンを生成しますが、血液中の溶解リン酸カルシウムのレベルが低いため、石灰化されません。 どちらの病気の症状にも、骨の変形や骨組織の全体的な軟化が含まれます。 もう 1 つの一般的な骨疾患は骨粗鬆症であり、これは高齢者によく発生します。 この病気では、骨物質の有機成分とミネラル成分の比率は変化しませんが、破骨細胞の活動の増加により、骨の形成よりも骨の吸収が激しくなるという事実が生じます。 骨粗鬆症の影響を受けた骨は徐々に薄くなり、もろくなり、骨折しやすくなります。 これらの結果は、脊椎骨粗鬆症で特に一般的です。
骨組織
骨組織 (textus ossei) は、細胞間有機物の高度な石灰化を伴う特殊なタイプの結合組織であり、約 70% の無機化合物、主にリン酸カルシウムを含んでいます。 骨組織には 30 種類以上の微量元素 (銅、ストロンチウム、亜鉛、バリウム、マグネシウムなど) が検出され、体内の代謝プロセスで重要な役割を果たしています。
骨組織の基質である有機物は、主にコラーゲン型のタンパク質と脂質で構成されています。 軟骨組織と比較して、それには比較的少量の水分、コンドロイチン硫酸が含まれていますが、カルシウムと複合体を形成するクエン酸やその他の酸が多く含まれており、骨の有機基質に浸透します。
したがって、骨組織の固体細胞間物質は(軟骨組織と比較して)骨に高い強度を与えると同時に、もろさを与えます。 有機成分と無機成分の相互作用により、骨組織の機械的特性、つまり張力と圧縮に耐える能力が決まります。
高度な石灰化にもかかわらず、骨組織は、構成物質の絶え間ない更新、絶え間ない破壊と生成、および変化する動作条件に対する適応的な変化を経験します。 骨組織の形態機能特性は、年齢、身体活動、栄養状態に応じて、また内分泌腺の活動、神経支配、その他の要因の影響を受けて変化します。
分類
骨組織には主に 2 つのタイプがあります。
網状繊維状(粗い繊維状)、
ラメラ状。
これらの種類の骨組織は、主に細胞間物質の構造によって決定される構造的および物理的特性が異なります。 粗い線維組織では、コラーゲン線維がさまざまな方向に走る太い束を形成し、層状組織では、骨物質(細胞、線維、基質)がプレートの系を形成します。
骨組織には、象牙質と歯科用セメントも含まれます。これらは、細胞間物質の高度な石灰化と支持する機械的機能の点で骨組織と似ています。
骨細胞: 骨芽細胞、骨細胞、破骨細胞。 それらはすべて、軟骨組織の細胞と同様、間葉から発生します。 より正確には、中胚葉強膜の間葉細胞から。 しかし、骨芽細胞と骨細胞は、線維芽細胞と線維細胞(または軟骨芽細胞と星状細胞)と同様に、分化して結合しています。 しかし、破骨細胞は血行性という異なる起源を持っています。
骨の分化と骨組織形成
胚における骨組織の発生は 2 つの方法で行われます。
1) 間葉から直接 - 直接骨形成。
2) 以前に開発された軟骨骨モデルの代わりに間葉から - これは間接的な骨形成です。
骨組織の胚後の発達は、その生理学的および修復再生中に起こります。
骨組織の発達中に、骨の差異が形成されます。
幹細胞、
半幹細胞(前骨芽細胞)、
骨芽細胞(線維芽細胞の一種)、
骨細胞。
2 番目の構造要素は、血液幹細胞から発生する破骨細胞 (マクロファージの一種) です。
幹細胞および半幹骨形成細胞は形態学的に同定されません。
骨芽細胞(ギリシャ語の osteon - 骨、blastos - 原始)は、骨組織を形成する若い細胞です。 骨では骨膜にのみ存在します。 それらは増殖する能力があります。 骨の形成において、骨芽細胞は、成長中の骨梁の表面全体をほぼ連続した層で覆っています。
骨芽細胞の形状は、立方体、錐体、または角張ったものなど、さまざまです。 体の大きさは約15〜20ミクロンです。 核は円形または楕円形で、多くの場合偏心して位置し、1 つまたは複数の核小体を含みます。 骨芽細胞の細胞質では、顆粒小胞体、ミトコンドリア、ゴルジ体がよく発達しています。 大量の RNA と高いアルカリホスファターゼ活性が含まれています。
骨細胞(ギリシャ語の osteon - 骨、cytus - 細胞に由来)は、分裂能力を失った骨組織の主な成熟(最終的な)細胞です。 それらは突起状で、コンパクトで比較的大きな核と弱好塩基性の細胞質を持っています。 オルガネラは十分に発達していません。 骨細胞における中心小体の存在は確立されていません。
骨細胞は、骨細胞の輪郭に沿った骨の隙間にあります。 キャビティの長さは 22 ~ 55 ミクロン、幅は 6 ~ 14 ミクロンの範囲です。 骨小管は組織液で満たされており、骨小管同士および骨に入る血管の血管周囲空間と吻合します。 骨細胞と血液の間の物質の交換は、これらの尿細管の組織液を通じて行われます。
破骨細胞(ギリシャ語のオステオン(骨とクラストス)が由来)は、石灰化した軟骨や骨を破壊する可能性がある造血性の細胞です。 直径は90ミクロン以上に達し、3個から数十個の核が含まれています。 細胞質はわずかに好塩基性であり、場合によっては好酸性です。 破骨細胞は通常、骨梁の表面に存在します。 破壊された表面に隣接する破骨細胞の側面には、細胞質突起(波形の境界)が豊富にあります。 それは加水分解酵素の合成と分泌の領域です。 破骨細胞の周囲には、細胞が骨表面にしっかりと接着しているゾーンがあり、いわば酵素の作用領域を密閉します。 細胞質のこのゾーンは軽く、アクチンからなるマイクロフィラメントを除いて細胞小器官はほとんど含まれていません。
波形の端の上の細胞質の周縁層には、多数の小さな小胞と大きな液胞が含まれています。
破骨細胞は環境中に CO2 を放出し、酵素炭酸脱水酵素は炭酸 (H2CO3) の生成とカルシウム化合物の溶解を促進すると考えられています。 破骨細胞にはミトコンドリアとリソソームが豊富に含まれており、その酵素(コラゲナーゼやその他のプロテアーゼ)は骨組織マトリックスのコラーゲンとプロテオグリカンを分解します。
1 つの破骨細胞は、100 個の骨芽細胞が同時に作成するのと同じ量の骨を破壊できると考えられています。 骨芽細胞と破骨細胞の機能は相互に関連しており、ホルモン、プロスタグランジン、機能的負荷、ビタミンなどによって調節されています。
細胞間物質(細胞間物質)は、無機塩が含浸された塩基性非晶質物質からなり、その中にコラーゲン線維が小さな束を形成して位置しています。 これらには主にタンパク質、I型およびV型コラーゲンが含まれています。 線維は、網状線維性骨組織ではランダムな方向を持ち、層状骨組織では厳密に配向された方向を持ちます。
骨組織の基底物質には、軟骨と比較して、比較的少量のコンドロイチン硫酸が含まれていますが、カルシウムと複合体を形成し、骨の有機基質に浸透するクエン酸やその他の酸が多く含まれています。 骨組織の主要物質には、コラーゲンタンパク質に加えて、非コラーゲン性タンパク質(オステオカルシン、シアロタンパク質、オステオネクチン、さまざまなリンタンパク質、石灰化プロセスに関与するプロテオリピド)、およびグリコサミノグリカンが見られます。 骨の基質には、骨の有機基質の原線維に関連して規則正しく配置されたヒドロキシアパタイトの結晶と、非晶質リン酸カルシウムが含まれています。 骨組織には 30 種類以上の微量元素 (銅、ストロンチウム、亜鉛、バリウム、マグネシウムなど) が検出され、体内の代謝プロセスで重要な役割を果たしています。 身体活動を体系的に増加させると、高度な石灰化により骨量が 10 ~ 50% 増加します。
骨骨格は 3 つの重要な機能を実行します。機械的、保護的、代謝的(代謝的)。 機械的機能。 骨、軟骨、筋肉は筋骨格系を形成しており、そのスムーズな動作は骨の強度に大きく依存します。 保護機能。骨は重要な器官(胸、頭蓋骨、骨盤、背骨)のフレームを形成します。 また、血球と免疫系の発達において重要な役割を果たす骨髄も収容されています。
代謝機能。骨組織はカルシウムとリンの貯蔵庫として機能し、その高い不安定性により体内のミネラル代謝に参加します。
海綿状の骨組織と緻密な骨組織があり、それらは同様の組成と基質構造を持っていますが、密度が異なります。
緻密な骨組織は成熟骨格の 80% を構成し、骨髄と海綿骨領域を取り囲んでいます。
緻密な骨組織と比較して、海綿骨組織は単位体積あたり約 20 倍の表面積を持っています。
緻密な骨と骨梁は骨髄の枠組みを形成します。
骨組織は ダイナミックシステム人の生涯を通じて、古い骨の破壊と新しい骨の形成のプロセスが発生し、これが骨組織の再構築のサイクルを構成します。 これは、骨が成長し、再生する一連の一連のプロセスです。
小児期および青年期には、骨は活発な再構築を受け、骨破壊(吸収)よりも骨形成が優勢になります。
骨は 2 つの主要な部分で構成されます。有機と無機。 骨の有機的基盤は、いくつかのクラスの細胞です。 骨芽細胞は構築細胞のグループを表し、破骨細胞は骨組織を破壊して余分なものを除去します。 骨の主な構造単位は骨細胞であり、コラーゲンを合成します。 骨細胞(骨芽細胞、骨細胞、破骨細胞)は骨の 2% を構成します。
骨細胞- 骨芽細胞由来の高度に分化した細胞。石灰化した骨基質に囲まれ、コラーゲン原線維で満たされた骨細胞腔に位置します。 成熟した人間の骨格では、骨細胞がすべての骨形成細胞の 90% を占めます。
骨芽細胞と骨細胞の生合成活性、およびこれに関連する細胞間物質の組織は、負荷ベクトルの大きさと方向、ホルモンの影響の性質と大きさ、および細胞の局所環境の要因に依存します。 したがって、骨組織は不安定で常に変化する構造です。
骨組織吸収の最も集中的な方法の 1 つは、 破骨細胞の吸収破骨細胞によって行われます。 それらはマクロファージ単球前駆体に由来する骨格外起源のものです。
骨基質体積の90%を占め、残りは細胞、血管、リンパ管で構成されています。 骨組織の細胞間物質は水分含有量が低いです。
骨基質は有機成分とミネラル成分で構成されています。 無機成分は骨の重量の約60%、有機成分は30%を占めます。 長期細胞と水は約 10% を占めます。 全体として、緻密骨中のミネラルマトリックスは、重量およびパーセンテージの点で有機マトリックスよりわずかに少ないです。
骨組織には、マグネシウム、銅、亜鉛、ストロンチウム、バリウムなどの 30 以上の微量元素が含まれており、これらは体内の代謝プロセスに積極的に関与します。
骨は体内の最大のミネラルバンクです。 カルシウム99%、リン85%、マグネシウム60%が含まれています。 ミネラルは体の必要に応じて常に消費されるため、補給する必要があります。
人生の特定の期間(妊娠、授乳中、子供の思春期、女性の更年期、ストレスの多い状況、腸や内分泌系の多くの病気、怪我によりカルシウムやビタミンの吸収が損なわれている場合)には、カルシウムの必要性が増加します。
特に カルシウムはホルモン変化の際にすぐに消費されてしまいます女性の体(妊娠、閉経)。 妊娠中の母親にとって、子供の骨格の正しい形成と発達、および将来の虫歯の有無はこれに依存するため、食品中の十分なカルシウム含有量に注意することが非常に重要です。 カルシウムの補給は、臓器やシステムの正常な機能だけでなく、骨粗鬆症を含む多くの病気の予防にも必要です。
通常、骨の合成と吸収のバランスは非常にゆっくりと変化します。 しかし、内分泌系(卵巣、甲状腺と副甲状腺、副腎のホルモン)と環境、その他多くの要因の両方から多くの影響を受けます。 そして、調節システムと代謝システムのわずかな機能不全は、ビルダー細胞とデストロイヤー細胞の間の不均衡を引き起こし、骨内のカルシウムレベルの低下を引き起こします。
ほとんどの人は、25 歳から 35 歳の間に最大骨量に達します。 これは、この時点で骨の密度と強度が最大になることを意味します。 残念ながら、これらの特性は将来的には徐々に失われ、骨粗鬆症の発症やその後の予期せぬ骨折につながる可能性があります。
骨組織の状態:
A - 通常。
B - 骨粗しょう症用
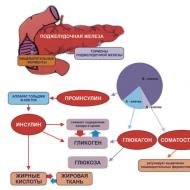
骨組織のモデリングとリモデリング一連の複雑な要因によってもたらされます。 これらには全身的要因が含まれており、そのうち 2 つのグループのホルモンを区別できます。
- カルシウム調節ホルモン(副甲状腺ホルモン、ビタミン03の活性代謝物であるカルシトリオール、カルシトニン);
- 他の全身ホルモン(糖質コルチコイド、性ホルモン、チロキシン、成長ホルモン、インスリンなど)。
インスリン様成長因子 (IGF-1、IGF-2)、線維芽細胞成長因子、トランスフォーミング成長因子 (TGF-β)、血小板由来成長因子など、大きなグループに結合した成長因子が骨リモデリングの制御に関与しています。要因など
骨代謝とミネラル代謝の調節における重要な役割プロスタグランジン、形態形成タンパク質、破骨細胞活性化因子など、細胞自体によって産生される他の微小環境因子も役割を果たします。
ホルモンの中で、骨代謝とカルシウムの恒常性に対する最も重大な影響は、副甲状腺ホルモン、ビタミン D およびその代謝産物によって及ぼされ、程度は低いですがカルシトニンによってもたらされます。 女性では、骨組織の代謝の調節はエストロゲンの影響を受けます。 体の腺によって生成される他のほとんどすべてのホルモンは、骨の再構築の調節に何らかの役割を果たしています。
骨および軟骨組織の前駆細胞
骨細胞間葉系(間葉系、中胚葉系)起源を持つ。 成人の体内では、骨と軟骨または骨髄組織との境界に局在する骨形成幹前駆細胞から形成されます。 分化すると、それらは骨芽細胞に変わり、次に骨細胞になります。 長い管状骨の成長は、軟骨内骨化によって起こります。 さらに、骨幹の幅の増加は骨膜からのみ発生し、骨幹端は骨内膜からのみ増加します。 したがって、骨吸収のプロセスは逆方向になります(Burne、1971、1976; Friedenstein、Lalykina、1973)。
A.Yaの作品に基づいて構築された、骨および軟骨組織の形成のスキーム。 フリーデンシュテイナ、E.A. ルリア (1980)、A.Ya。 フリーデンシュタインら (1999)、I.L. チェルトコバ、O.A. グレヴィッチ (1984)、VP。 シャホワ (1996)。 N. Castro-Malaspina et al. (1980, 1982) にいくつかの修正を加えたものを図に示します。
骨形成、軟骨形成、破骨細胞形成のスキーム。 CKKH - 骨および軟骨組織の幹細胞、CKK - 造血幹細胞、PPKK - 造血組織の多能性前駆細胞、PCKH - 骨および軟骨組織の多能性前駆細胞、B(U)KPKH - 二(一)能性前駆細胞骨および軟骨組織、KPKM - 造血微小環境を運ぶ細胞、CFUf - 線維芽細胞のコロニー形成単位、U (B) KPK (X、M、G、E、Meg、T、V) - 単能性 (双能性) 前駆体骨の細胞(軟骨、マクロファージ、顆粒球、赤血球、巨核球、TおよびBリンパ球)組織
骨組織形成のプロセスは、複雑な多段階のプロセスであり、さまざまな組織遺伝学的系統の細胞が増殖、分化、特殊化を通じて連続的に変換を受けて、骨と呼ばれる複合構造を形成します。
骨および軟骨組織が胚発生において中胚葉の背側体節から形成される場合、破骨細胞の起源となる造血組織は内臓中胚葉の段階を経ることが強調されるべきである。 組織形成の観点から見ると、骨細胞と骨芽細胞は結合組織、筋肉、皮膚の要素に近く、破骨細胞は血球や内皮に近い(Coalson、1987)。 破骨細胞芽腫における上皮組織と筋肉組織の存在は、この見解を裏付けるようです。
胚発生における造血から骨軟骨形成の発生の方向が分岐した後、成熟した生物では、より分化して組織に固定された、または循環する未熟な間質要素(中胚葉細胞、未分化線維芽細胞、骨形成前駆体または前駆体)(Friedenstein、Luria、1980; Alberst et al.、1994; Omelyanchenko et al.、1997)。 骨および軟骨組織の多能性幹細胞の存在に加えて、より分化した前駆体も存在します。 BMSC は高い増殖能を持ち、多能性を持っています。 それらは、少なくとも骨および(または)軟骨核細胞を形成し、これらは主に細胞周期の G1 ~ G2 段階にあります(Fridenstein、Lalykina、1977; Friedenstein、Luria、1980; Friedenstein et al.、1999; Chertkov、グレビッチ、1984 年)。
インビボおよびインビトロの組織培養において、それらは軟骨組織または骨組織を形成し、これらは線維芽細胞-COEFのコロニー形成単位と呼ばれるコロニーの形態で存在することができる(FriedensteinおよびLuria、1980)。 放射線キメラの染色体および生化学マーカーを使用して、CFU が骨芽細胞や骨細胞を含む骨髄の造血細胞とは起源が異なるクローン性を有することが示されました (Chertkov および Gurevich、1984)。
我々は、培地に導入された核球の数と、Balb/c マウスの骨髄組織の浮遊培養で形成されるコロニーの数との関係を研究しました。 これを行うために、骨髄をシリコン処理したチューブに洗浄し、20%ウシ胎児血清、40μg/mlゲンタマイシン、200mM L-グルタミンhepesを含むD-MEM培地に懸濁し、プラスチック中で2〜3週間培養しました。バイアルは37°で。 播種密度は 1 ml あたり 104 ~ 107 細胞の範囲でした。
Balb/c マウスのさまざまな量の骨髄細胞を培養物に導入したときの CFU 形成の依存性
提示されたデータは、一般に、培養物に導入された骨髄核球の数とCFUfの間の関係が線形であることを示しており、これは再びそれらのクローン起源を裏付けています。
腎臓被膜の下または皮膚の下に移植すると、骨または軟骨組織を形成する能力があります。

ストレスを受けた F1 マウスからの骨髄移植後に腎臓被膜の下で成長した異所性骨組織の肉眼標本 (CBAxC57Bl)。 左側、臓器の上部極に、骨形成の大きな焦点がはっきりと見えます。 右側 - 対照(ストレスを受けていない動物から採取した骨髄)
BMSC の特性の 1 つは、元の培養物をあるドナーから別のドナーに繰り返し移植しても、BMSC の増殖および分化の潜在能力が保持されることです。 どうやら、このレベルのゲノム損傷は骨肉腫の形成につながるようです。
BMSC の分化の結果として、より分化した BPKC タイプの前駆細胞 (骨および軟骨組織の前駆細胞) または BKKKH (両能性) が形成され、次に UPKKH および UPKKH (骨または軟骨組織に対する単能性) が形成されます。 骨を含むあらゆる組織の前駆細胞のプールの一般的なパターンは、自己複製および増殖能力の段階的な低下、多能性の喪失、細胞の S 期に位置する前駆細胞の割合の増加です。成長因子、ホルモン、サイトカイン、その他の調節分子の作用に対する感受性の増加。 理論的には、このプロセスは均一に進行することも、断続的に進行することもあります。 このため、骨形成の過程は、形態機能特性において質的および量的に異なる骨組織の形成とともに、さまざまなモードおよび速度で進行する可能性があります。 私たちの意見では、骨への生体材料の導入には、必ず何らかの骨形成細胞の発達経路が含まれるでしょう。 しかし、残念ながら、この非常に興味深い方向性で行われた研究は見つかりませんでした。
PKPKH が多分化能を持つ場合、BKPKH は軟骨または骨組織を形成し、UKPKH は骨のみを形成し、UKPKH は軟骨を形成します。 前駆細胞のすべてのカテゴリーは非常に不均一な集団を表しており、その中で形態機能特性が広範囲にわたって変化することに留意すべきである。 さらに、CP 開発の各段階には、既存のテクノロジーを使用してもまだ特定できない移行形態が多数存在します。 間質前駆細胞と骨形成前駆細胞を同定する方法が 70 年代初頭に発見されたという事実にもかかわらず、それらの特性、調節方法、骨組織再構築のプロセスにおける役割の理解は明らかな進歩を遂げていません (Fridenstein、Lalykina、1973; Friedenstein ら、1999; Chertkov、Gurevich、1984; Setsulla、Devyatov、1987; Omelyanchenko ら、1997)。
骨および軟骨組織の幹細胞および分化された前駆細胞は、局所的および遠隔の調節機構の制御下にあることに留意すべきである。 最後のグループには、長距離メッセンジャー (エストロゲン、グルココルチコイド、エンドルフィン、アドレナリンなど) を生成または結合することによって、神経内分泌、免疫、細網内皮、アヘン剤、NO およびその他のシステムを通じて効果を発揮する因子が含まれます。 局所メカニズムは、骨組織の微小環境、細胞間接触、サイトカイン、メディエーター、短寿命の生理活性物質などの局所的生成の形態機能特性の直接的な変化を通じて機能します。 細胞間相互作用は形態形成プロセスに属し、組織や器官における細胞の分化、特殊化、および形態形成を制御します。 それらの実装メカニズムは、位置情報と帰納的相互作用を使用して実行されます。 それらはまだほとんど研究されていません。 しかし、位置情報の概念によれば、身体には形態形成場が存在する。 それは、HOX1、HOX2、HOX3、HOX4、HOX7 などのホメオティック遺伝子の発現によって制御されており、細胞は座標軸に従ってその局在の場所を記憶するだけでなく、細胞が果たさなければならない使命を遂行することも強制されます。たとえば、骨が損傷した場合は修復するなど、生涯にわたって行う必要があります。 間葉系要素、特にマクロファージ、骨芽細胞、骨細胞、破骨細胞、内皮および線維芽細胞は、位置情報の維持において重要な役割を果たしていると考えられている(Gilbert、1994)。
誘導機構は、サイトカイン、成長因子、さまざまな代謝産物、短距離メッセンジャーの助けを借りて、直接的な細胞相互作用に至るまで、自己複製細胞集団の増殖と分化のプロセスを制御します。
多能性および二能性骨形成前駆体の分化方向の選択の特徴は、主に酸素分圧に依存することです。 この圧力が十分に高い場合、骨前駆体は骨形成の方向に発達し、圧力が低い場合は逆に、軟骨組織を形成します(Bassett、Herman、1961)。 細胞への適切な酸素供給は、発達した微小血管網の存在下でのみ可能であることを覚えておく必要があります。骨前駆体の除去の最大量は 100 μm を超えてはなりません (Ham and Cormack、1983)。
オステオンシステム
成人の骨のハバース システムは常に更新されます。 この場合、進化中または発達中(5-10%)、成熟中(50-75%)、変性中または退縮中(10-20%)、再構築中(5-10%)など、いくつかのタイプの骨を区別することが常に可能です。および生存不能 (5 ~ 10).%)。
オステオン(Haversian system)は、単球、マクロファージ、および破骨細胞の作用の結果として形成されたトンネルに基づいてのみ発生し、骨芽細胞と破骨細胞によって形成された骨組織の濃縮層で内側から満たされていると考えられています(ハムおよびコーマック) 、1983)。 オステオンシステムは常に進化する可動構造であることに注意してください。 逆説的ですが、骨動態の研究に特化した研究はほとんどありません。 放射性核種の研究方法を使用すると、骨組織の表層の年間置換率は 5 ~ 10% であることがわかりました (Harris、Heaney、1969)。 どうやら、オステオンの更新速度にも同様のパラメーターがあるようです。 興味深いことに、発生中のオステオンの直径は一定の値ではなく、生涯を通じて何度も連続的に変化します。 文献と私たち自身のデータの分析により、セメント結合線によって制限されるハバースシステムの境界は、若い、発達中および再構築中の骨骨では80〜150μm、成熟した骨骨では120〜300μm、退縮期では120〜300μmであると信じることができます。 、変性 - 200 μm未満。 骨形成のプロセスが骨膜と骨の境界で発生する場合、チャネルの代わりに溝が最初に形成され、その壁には増殖する骨形成細胞が並んで隆起を形成します。 これらの細胞突起の壁は互いに接近して空洞を形成し、その中には通常、少なくとも 1 本の栄養動脈が存在します。 次に、骨形成細胞は骨芽細胞と骨細胞に分化し、骨を形成します。 外傷学で使用される材料は、骨のサイズと等しい孔径を有するべきであることが示唆されている(Gunther et al.、1992)。 しかし、これらの著者らは、孔径が発生、再構築、成熟した骨の直径に対応するべきであるという主な基準を実証しなかった。 この原理が孔径を大きくしたり小さくしたりする方向に破られると、本格的な骨組織は形成されなくなります。 つまり、骨の大きさは人工骨組織を作成する際に考慮すべき重要な形態形成因子であると考えられます。 この現象のメカニズムは完全には明らかではありません。 それはおそらく骨形成細胞自体に遺伝的にプログラムされており、骨の微小環境の重要な要素です。 同時に、材料を作成する際には、骨の直径などの体積特性に加えて、以下で説明する他の生物学的原理を考慮する必要があることを強調する必要があります。
AV カルポフ、VP シャホフ
硬骨魚類には、魚座のスーパークラス全体の大部分 (20,000 以上) の種が含まれます。 硬骨魚はさまざまな水域でよく見られます。 生活条件の多様性が、このグループの種の豊かさとその極端な多様性を決定します。
硬骨魚綱にはすべての硬骨魚が含まれます。 スケール - 形状に応じてサイクロイドまたはクテノイド - それぞれ滑らかまたは鋸歯状。 種の数と形態の多様性の点で、硬骨魚は軟骨魚よりもはるかに優れています。 おそらく最も進化しているのは、ニシン、マス、サケ、コイ、ウナギ、トビウオなどを含むテレオステイ目(硬骨魚)です。
クラスの主な一般的な特徴は次のとおりです。
骨格は常に多かれ少なかれ骨化しています。 骨の骨格は 2 つの方法で生成されます。 最初のタイプの骨化は、いわゆる皮膚、または外皮の骨です。 胚性の場合、それらは、それらが隣接するだけの骨格の軟骨要素に関係なく、皮膚の結合組織層で発生します。 これらの発達上の特徴により、外皮骨は通常、プレートの形をしています。 外皮骨に加えて、魚の骨格には軟骨、つまり軟骨の骨が含まれています。 胚性のそれらは、骨芽細胞によって生成される骨物質による軟骨の連続的な置換の結果として発生します。 組織学的に形成された軟骨骨は、外皮骨と大きな違いはありません。 軟骨骨の出現によって起こる骨格の骨化は、骨格の全体的な構造に大きな変化をもたらしません。 外皮骨化の形成は、新しい骨格要素の出現をもたらし、その結果、一般的な合併症を引き起こします。
呼吸器の枝間隔壁は減少し、鰓糸は鰓弓に直接位置します。 鰓装置の外側を覆う骨蓋が常に存在します。
ほとんどの種には浮き袋があります。
硬骨魚の大部分では体外受精が行われ、卵は小さく、角のような膜がありません。 胎生はごく少数の種で起こります。 硬骨魚類の分類は非常に難しく、現在、このグループの分類法についてはいくつかの見解があります。 そのうちの 1 つを基礎として、2 つのサブクラスを区別します。
1) サブクラス 条鰭魚 (Actinopterygii) 2) 亜綱 条鰭魚 (Sarcopterygii)。
硬骨魚の外部構造と内部構造。
外部構造
体の大きさは1cm(フィリピンハゼ)から17m(ニシンキング)まであります。 ブルーカジキの体重は最大900kgです。 体の形は通常、細長く流線形ですが、硬骨魚の中には背腹側または横側が平らになっているものや、その逆の球形のものもいます。 水中での並進運動は、体の波のような動きによって行われます。 一部の魚は尾鰭で自分自身を「助ける」。 一対の横鰭、背鰭、臀鰭は安定舵として機能します。 一部の魚では、個々のヒレが吸盤または交尾器官に変化しています。 硬骨魚の体の外側は、プラコイド(「寄木細工」に置かれた歯)、ガノイド(棘のある菱形の板)、サイクロイド(滑らかな端を持つ薄い板)、またはクテノイド(棘のある板)といった鱗で覆われています。動物の成長に合わせて定期的に交換されます。 年輪によって魚の年齢が判断できます。 さまざまな種類の鱗 多くの魚の皮膚にはよく発達した粘液腺があり、その分泌物によって水の流れに対する抵抗力が軽減されます。 一部の深海魚は皮膚に発光器官を発達させ、種を識別し、群れを強化し、獲物をおびき寄せ、捕食者を追い払うのに役立ちます。 これらの器官の中で最も複雑なものはスポットライトに似ています。これらの器官には、発光要素 (燐光バクテリアなど)、鏡の反射板、絞りまたはレンズ、および黒または赤の絶縁コーティングが備わっています。 魚の体色は非常に多様です。 通常、魚は青みがかったまたは緑がかった背中(水の色と一致する)、銀色の側面と腹(明るい「空」を背景にするとほとんど見えません)を持っています。 多くの魚は迷彩のために縞模様や斑点で覆われています。 それどころか、サンゴ礁の住民は色彩の暴動に驚かされます。
消化器系
食物は口腔から咽頭に入り、そこから食道に入り、次に大きな胃に入るか、直接腸(フナ)に入ります。 胃では、胃液の影響下で食物の部分消化が起こります。 食物の最終的な消化は小腸で行われます。 胆嚢管、肝臓管、膵管は小腸の最初の部分に流れ込みます。 小腸では、栄養素が血液中に吸収され、未消化の食べ物の残りが肛門から除去されます。
排泄系
水生環境で生活すると、淡水魚と海水魚の両方に影響を与える多くの浸透圧調節の問題が発生します。 魚の血液の浸透圧は、外部環境の浸透圧よりも低い場合(海水魚の場合)と高い場合(淡水魚の場合)があります。 軟骨魚は等浸透圧ですが、同時に体内の塩分濃度は環境よりもはるかに低くなります。 この場合、血液中の尿素とトリメチルアミンオキシド(TMAO)の含有量が増加することにより、浸透圧の均一化が達成されます。 軟骨魚の体内の塩分濃度を低く維持できるのは、腎臓と、消化管につながる特殊な直腸腺による塩分の分泌によるものです。 直腸腺は、血液および身体組織からナトリウムイオンと塩化物イオンの両方を濃縮して除去します。 硬骨魚類は等張性ではないため、進化の過程でイオンの除去または保持を可能にする機構を発達させてきました。 体内のイオン濃度が(環境に比べて)低い海洋硬骨魚は常に水分を失い、浸透圧の影響で組織が外部に出てしまいます。 これらの損失は、塩水を飲んだり濾過したりすることで補われます。 ナトリウム陽イオンと塩化物イオンは鰓膜を通って血液から除去され、マグネシウム陽イオンと硫酸陰イオンは腎臓から排泄されます。 淡水魚は逆の問題に直面しています(環境よりも体内の塩分濃度が高いため)。 鰓膜を通した水生環境からのイオンの捕捉と、大量の尿素の放出により、それらの浸透圧は均一化されます。
呼吸器系
えらを通して呼吸します。 口腔からは鰓裂を通って水が鰓を洗い、鰓蓋の下から出てきます。 鰓は鰓弓で構成され、鰓弓は鰓糸と鰓かきで構成されています。 一部の種では、皮膚呼吸が不可欠であるか、空気呼吸への適応が存在します。
循環系
魚の循環系は閉じており、心臓は心房と心室の 2 つの部屋で構成されています。 大きな血管である大動脈は心室からえらまで伸び、小さな血管である動脈に分岐します。 えらでは、動脈は小さな血管の密なネットワーク、つまり毛細血管を形成します。 血液が酸素を豊富に含むと(酸素が豊富な血液は動脈血と呼ばれます)、血管は再び集合して動脈になり、さらに小さな動脈と毛細血管に分岐します。 体の器官では、毛細血管の壁を通って酸素と栄養素が組織に入り、組織から二酸化炭素やその他の老廃物が血液に入ります。
繁殖システム
ほとんどの魚の生殖腺は対になっていて、浮き袋のすぐ下の後ろの体腔にあります。 しかし、メスのカワスズキでは、卵巣は不対になります。 生殖器の開口部は、肛門開口部と排泄口の間の体の後部に開きます。 大部分の魚では、生殖細胞が外部環境に放出される体外受精が観察されます。 繁殖期における魚の複雑な本能的な行動は、産卵と呼ばれます。 リバーパーチは生後2年目に成熟します。 水域の氷がなくなるとすぐに産卵し始めます。 産卵前は特に魚の色が明るくなります。 流れの弱い場所に大きな群れを作って集まります。 メスは接着された長いリボンの形で卵を産み、水中の植物に定着します。 同時に、これらの雄は数百万の小さな精子を含む乳を生成します。
浮き袋
スズキの内腔の上部、腸の上には、ガスで満たされた半透明の袋のように見える、ボリュームのある浮き袋があります。 魚は水より重いため、その主な機能は魚にプラスの浮力を与えることです。 浮き袋は密な血管網に囲まれており、そこからガスが放出されます。 浮き袋の容積が増加すると、スズキの体の密度が減少し、水面に浮かび上がります。 逆に体積が減ると身密度が増し、魚は底に沈んでしまいます。 浮き袋は、腸の背壁の増殖物として発達します。 魚は特別な静水器官、側線を発達させました。 頭から尾まで体に沿って伸びる一連の小さな毛穴のように見えます。 毛穴は皮膚にある管に通じています。 多数の神経終末がそこに接近します。 側線の助けを借りて、魚は水流の方向と強さ、浸水の深さ、さまざまな物体の接近を評価します。 鰓蓋には水圧を感知する器官が発達しているため、側線はニシン目の代表者にのみ存在します。
硬骨魚の骨格。
硬骨魚では、骨格内の軟骨がある程度骨組織に置き換えられ、主骨または代替骨が形成されます。 さらに、外皮骨が皮膚に出現し、皮膚の下に沈んで内部骨格の一部を形成します。 硬骨魚の骨格は、軸骨格、頭蓋骨(脳および内臓)、不対のヒレの骨格、対のヒレの骨格およびそれらのベルトに分けられます。
葉鰭、肺魚、およびチョウザメに似た魚では、軸骨格における支持機能は、高密度の結合組織膜に囲まれた脊索によって行われます。 よく発達した、時には部分的に骨化した上弓は、脊髄が存在する管を形成します。 肋骨は、発達が不十分な下部アーチに付着しています。 これらのグループの祖先の中には、多かれ少なかれ発達した椎体を持った人もいました。 羽毛の多い魚やすべての硬骨魚では、両親媒性タイプ (両凹面) の骨椎骨がよく発達しています。 強く縮小した脊索は独特の構造を持っています。脊索は椎体間の空間で拡張され、強く狭まった形で椎体の中心の管を通過します。 幹の椎骨には、長い上棘突起で終わる骨の上アーチがあります。 長くて薄い骨質の肋骨は、ほとんどの硬骨魚でよく発達しており、椎体の横突起に取り付けられています。 尾部の椎骨には棘突起のある上部アーチがあり、横突起は下方に移動し、対になって結合して下部アーチを形成し、下部突起で閉じられています。 アーチ間の空間は緻密な結合組織フィルムで覆われています。 上管には脊髄が含まれています。 尾椎の下部アーチは尾動脈と尾静脈が通る血液管を形成し、この部分の強力な筋肉による圧迫から保護されています。 椎骨は、上部アーチの基部にある関節突起を使用して互いに接続されています。 このような関節は、軸骨格の可動性を維持しながら、軸骨格に強度を与えます。 脊柱は主に水平面内で曲がることがあります。 ほとんどの硬骨魚では、薄い筋骨が筋肉の深部にあり、筋線維をさらにサポートしています。
すべての脊椎動物と同様、硬骨魚の頭蓋骨は髄質 (軸方向) 部分と内臓部分に分かれています。
チョウザメでは、脳の頭蓋骨は軟骨のままです。 古い魚にのみ小さな骨化が現れます。 外側では、軟骨性の頭蓋骨は多数の外皮骨の連続した殻で覆われています。 しかし、ほとんどの硬骨魚は、脳の頭蓋骨の軟骨が、互いに直接隣接するか、または軟骨の残存物によって接続された主骨と、比較的少数の外皮骨(主な殻の残存物)によって置換されることを特徴としています。 脳頭蓋骨の骨化は、ガノイド(チョウザメを除く)と硬骨魚で高度に発達しています。 葉鰭類および肺魚類では、大量の軟骨が頭蓋骨に保持され、主要な骨は少数しか発達しません。 外皮骨の一次殻はよく発達しています。
硬骨魚では、後頭部分に大後頭孔に隣接して 4 つの骨、つまり主後頭骨 (基後頭骨)、側方の 2 つの骨 (外側後頭骨)、および上部後頭骨 (上後頭骨) が形成されます。 頭蓋骨の側壁には 5 つの耳の骨 (ossa otici) があります。 眼窩の領域では、蝶形骨が発生します:眼蝶形骨、基部蝶形骨(基底蝶形骨)、および外側蝶形骨(外側蝶形骨)。 嗅覚領域の領域では、対になっていない中骨(メセトモイデウム)と対をなす側方嗅骨(エクトエモイデウム)が形成されます。 これらの骨はすべて基本的な骨であり、軟骨領域の骨化によって発達します。 頭蓋骨の上部は、鼻骨(鼻骨)、非常に大きな前頭骨(フロンターレ)、小さな頭頂骨(頭頂骨)の3対の外皮骨で覆われています。 頭蓋骨の底部は、2 つの対になっていない外皮骨、すなわち大きな傍蝶形骨 (parasphenoideutn) と歯が生えている鋤骨 (votner) によって形成されています。 骨の外側のレリーフは複雑であるため、骨の間の境界は常に見えるわけではありません。
頭蓋骨の内臓骨格は、上顎骨、舌骨、および5つのえらなど、さまざまな程度の軟骨または骨化した弓のシステムです。 4 つの外皮骨が鰓蓋を形成します。 外皮骨は顎弓を強化し、二次顎を形成します。 硬骨魚は、舌骨弓、つまり舌骨弓の上部要素であるペンダントまたは顎顎骨を介して顎弓および副顎と脳の頭蓋骨との接続を特徴としています。 肺魚類でのみ、上顎が頭蓋骨の底部と融合し(自己固着)、機能を失った振り子はサイズが小さくなります。
チョウザメの内臓骨格には多くの軟骨が残存しており、第二顎はあまり発達していません。 硬骨魚では、この部分の軟骨が完全に骨に置き換わります。
主な上顎の口蓋方形軟骨の骨化の結果として、歯のある口蓋骨(口蓋骨)が両側に形成され、後部には後翼突骨(中翼突骨)と方形骨(方形骨)が形成されます。 それらの間には、外皮の外側翼突骨と内側翼突骨 (外翼突骨と内翼突骨) があります。 主な下顎 - 骨化したメッケル軟骨は関節骨 (articulare) に変化し、方形骨と顎関節を形成します。 硬骨魚は外皮骨から作られたよく発達した第二の顎を持っています。 それらは一緒に成長するか、靱帯によって主顎の骨に接続されます。 上顎では、そのような二次構造は前上顎骨 (preemaxillare) と上顎骨 (maxillare) になります。 歯はそれらの上にあります(いくつかの種では、歯は一方または両方の骨にありません)。 強力な外皮歯骨 (歯骨) が下顎の主要部分を形成しています。 獲物を掴んで保持する機能は、主顎と副顎の両方によって実行されます。
歯の力と特徴、顎の相対的なサイズと口の開口部の位置は、それぞれの種の摂食の専門性を反映しています。
舌骨弓は主要な骨によって形成されます。 その上部要素であるペンダントは、軸方向の頭蓋骨の聴覚部分に取り付けられた広い上端を備えた大きな顎骨で表されます。 ペンダントの下端から分離した追加の骨 symplecticum hyomandibulare を介して方形骨 (舌骨柱!) に取り付けられ、短い骨化した硝子体間靱帯を介して舌骨弓の下部要素である舌骨に取り付けられています。 、そこではいくつかの骨化が形成され、多くの場合、総骨の舌骨に融合します。 左右の舌骨の前端は、舌のひだを支える不対の骨である結合骨によって互いに接続されています。 薄い湾曲した骨が鰓膜の舌骨に取り付けられ、鰓蓋の革のような端を支えています。 硬骨魚類の鰓蓋は外皮骨によって形成されます。 強力で鋭く湾曲した鰓前骨 (鰓前骨) がペンダントおよび方形骨の後端にしっかりと隣接しており、鰓蓋骨、鰓蓋間骨、および鰓下骨が取り付けられています。
鰓弓は5対。 最初の 4 つは、関節によって互いに接続された 4 つの骨化した対の要素で構成されます。 その下にある 5 番目の対になっていない要素は、円弧を互いに接続します。 これらの鰓弓には鰓が付いています。 5 番目 (後部) の鰓弓は、2 つの大きな対になった要素のみで構成されます。 いくつかの種では咽頭歯があります(咽頭歯の形と大きさは摂食の特殊性を反映しています)
対になった手足とそのベルト。 対になった手足は胸鰭と腹鰭で表されます。 胸鰭は肩帯によって魚の体内で支えられています(図39)。 それは、2つの小さな置換骨(一次)といくつかの外皮骨によって表されます。 置換骨の上部 - 肩甲骨(肩甲骨、図39、1) - は、自由肢の関節領域にあります(骨の中心にある小さな丸い穴によって簡単に区別できます)。 そのすぐ下にはカラス骨、または烏口骨(烏口骨、烏口骨、図39、2)があります。 これら 2 つの要素が主ベルトを構成します。 それらは大きな外皮骨の頭蓋骨に動かずに接続されており(図39、3)、その上端はわずかに前方を向いています。 それは小さな骨である頭蓋上によって結合されています;図 39、4。
次に、頭蓋骨は後頭頂骨に接続します。 左右の頭頂部の前方を向いた下端は互いに接続されている。 鎖骨の後ろ、肩甲骨と烏口骨からそれほど遠くないところに、小さな後鎖骨があります(鎖骨後、図39、6)。 名前付きボーンはすべてペアになっています。 それらは二次肩帯を構成します。 左右の後頭頂骨は軸頭蓋骨に取り付けられており、ガードルの固定が強化され、その支持機能が強化されています。
胸鰭の基部には、肩甲骨から(部分的に烏口骨から)伸びる放射状の小さな骨が1列あります。 自由鰭の刃全体は、セグメント化された皮膚光線で構成されています1 (鱗翅目、図 39、8)。 軟骨魚と比較した硬骨魚の胸鰭の骨格の特徴は、基底骨が減少していることです。 胸鰭の可動性は、筋肉が皮膚線の拡張した基部に取り付けられており、橈骨と可動に関節運動するため、増加します。
骨盤帯は、互いに融合した一対の平らな三角形の骨によって表され、腹筋の厚さの中に位置し、軸骨格とは接続されていません。 骨盤鰭は骨盤帯の側面に取り付けられています。 ほとんどの硬骨魚類では、腹鰭の骨格には基底部がなく、橈骨が完全に縮小しています。鰭の刃は皮骨条(鱗翅条)によって支えられており、その拡張した基部は骨盤帯に直接取り付けられています。 腹鰭の骨格がこのように単純化されているのは、明らかにその機能が限定されているためです。
対になっていない手足。 対になっていない四肢は、背鰭、尾鰭下 (肛門) および尾鰭で表されます。 肛門鰭と背鰭は骨条で構成されており、内部(筋肉の厚さに隠れている)翼状翅と外部鰭条 - 鱗翅条に分かれています。
骨組織
骨組織 ( テクストゥス・オッセイ) は特殊なタイプの結合組織であり、 高い石灰化約70%の無機化合物を含む細胞間有機物、主にリン酸カルシウム。 骨組織には 30 種類以上の微量元素 (銅、ストロンチウム、亜鉛、バリウム、マグネシウムなど) が検出され、体内の代謝プロセスで重要な役割を果たしています。
骨組織の基質である有機物は、主にコラーゲン型のタンパク質と脂質で構成されています。 軟骨組織と比較して、それには比較的少量の水分、コンドロイチン硫酸が含まれていますが、カルシウムと複合体を形成するクエン酸やその他の酸が多く含まれており、骨の有機基質に浸透します。
したがって、骨組織の固体細胞間物質は(軟骨組織と比較して)骨に高い強度を与えると同時に、もろさを与えます。 有機成分と無機成分の相互作用により、骨組織の機械的特性、つまり張力と圧縮に耐える能力が決まります。
高度な石灰化にもかかわらず、骨組織は、構成物質の絶え間ない更新、絶え間ない破壊と生成、および変化する動作条件に対する適応的な変化を経験します。 骨組織の形態機能特性は、年齢、身体活動、栄養状態に応じて、また内分泌腺の活動、神経支配、その他の要因の影響を受けて変化します。
分類
骨組織には主に 2 つのタイプがあります。
網状繊維状(粗い繊維状)、
· ラメラ。
これらの種類の骨組織は、主に細胞間物質の構造によって決定される構造的および物理的特性が異なります。 粗い線維組織では、コラーゲン線維がさまざまな方向に走る太い束を形成し、層状組織では、骨物質(細胞、線維、基質)がプレートの系を形成します。
骨組織には、象牙質と歯科用セメントも含まれます。これらは、細胞間物質の高度な石灰化と支持する機械的機能の点で骨組織と似ています。
骨細胞: 骨芽細胞、骨細胞、破骨細胞。 それらはすべて、軟骨組織の細胞と同様、間葉から発生します。 より正確には、中胚葉の強核の間葉細胞から。 しかし、骨芽細胞と骨細胞は、線維芽細胞と線維細胞(または軟骨芽細胞と星状細胞)と同様に、分化して結合しています。 しかし、破骨細胞は血行性という異なる起源を持っています。
骨の分化と骨組織形成
胚における骨組織の発生は 2 つの方法で行われます。
1) 間葉から直接 - 直接骨形成。
2) 以前に開発された軟骨骨モデルの代わりに間葉から - これは間接的な骨形成です。
骨組織の胚後の発達は、その生理学的および修復再生中に起こります。
骨組織の発達中に、骨の差異が形成されます。
・ 幹細胞、
半幹細胞(前骨芽細胞)、
骨芽細胞(線維芽細胞の一種)、
· 骨細胞。
2 番目の構造要素は、血液幹細胞から発生する破骨細胞 (マクロファージの一種) です。
幹細胞および半幹骨形成細胞は形態学的に同定されません。
骨芽細胞(ギリシャ語より オステオン-- 骨、 爆風- 初歩)、骨組織を形成する若い細胞です。 骨では骨膜にのみ存在します。 それらは増殖する能力があります。 骨の形成において、骨芽細胞は、成長中の骨梁の表面全体をほぼ連続した層で覆っています。
骨芽細胞の形状は、立方体、錐体、または角張ったものなど、さまざまです。 体の大きさは約15〜20ミクロンです。 核は円形または楕円形で、多くの場合偏心して位置し、1 つまたは複数の核小体を含みます。 骨芽細胞の細胞質では、顆粒小胞体、ミトコンドリア、ゴルジ体がよく発達しています。 大量の RNA と高いアルカリホスファターゼ活性が含まれています。
骨細胞(付録の図 4、5 を参照)(ギリシャ語より オステオン-- 骨、 サイトス-- 細胞) は、分裂能力を失った骨組織の主な成熟 (最終的な) 細胞です。 それらは突起状で、コンパクトで比較的大きな核と弱好塩基性の細胞質を持っています。 オルガネラは十分に発達していません。 骨細胞における中心小体の存在は確立されていません。
骨細胞は嘘をつく 骨の隙間にある、骨細胞の輪郭に従います。 キャビティの長さは 22 ~ 55 ミクロン、幅は 6 ~ 14 ミクロンの範囲です。 尿細管骨の空隙は組織液で満たされており、互いに吻合したり、骨に入る血管の血管周囲の空間と吻合したりしています。 骨細胞と血液の間の物質の交換は、これらの尿細管の組織液を通じて行われます。
破骨細胞(ギリシャ語より オステオン-- 骨と クラストス- 破砕)は、石灰化した軟骨や骨を破壊する可能性がある血行性の細胞です。 直径は90ミクロン以上に達し、3個から数十個の核が含まれています。 細胞質はわずかに好塩基性であり、場合によっては好酸性です。 破骨細胞は通常、骨梁の表面に存在します。 破壊された表面に隣接する破骨細胞の側面には、細胞質突起が豊富にあります( 波形の境界線); それは加水分解酵素の合成と分泌の領域です。 破骨細胞の周囲には、 タイトシールゾーン細胞を骨表面に付着させ、いわば酵素の作用領域を密閉します。 細胞質のこのゾーンは軽く、アクチンからなるマイクロフィラメントを除いて細胞小器官はほとんど含まれていません。
波形の端の上の細胞質の周縁層には、多数の小さな小胞と大きな液胞が含まれています。
破骨細胞は環境中に CO 2 を放出し、酵素は 炭酸脱水酵素炭酸(H 2 CO 3 )の生成とカルシウム化合物の溶解を促進します。 破骨細胞にはミトコンドリアとリソソームが豊富に含まれており、その酵素(コラゲナーゼやその他のプロテアーゼ)は骨組織マトリックスのコラーゲンとプロテオグリカンを分解します。
1 つの破骨細胞は、100 個の骨芽細胞が同時に作成するのと同じ量の骨を破壊できると考えられています。 骨芽細胞と破骨細胞の機能は相互に関連しており、ホルモン、プロスタグランジン、機能的負荷、ビタミンなどによって調節されています。
細胞間物質 (細胞間質)は、無機塩が含浸された塩基性非晶質物質から構成されており、その中にコラーゲン線維が位置し、小さな束を形成しています。 主にタンパク質が含まれています - コラーゲンタイプIおよびV。 線維は、網状線維性骨組織ではランダムな方向を持ち、層状骨組織では厳密に配向された方向を持ちます。
骨組織の基底物質には、軟骨と比較して、比較的少量のコンドロイチン硫酸が含まれていますが、カルシウムと複合体を形成し、骨の有機基質に浸透するクエン酸やその他の酸が多く含まれています。 骨組織の主要物質には、コラーゲンタンパク質に加えて、非コラーゲン性タンパク質(オステオカルシン、シアロタンパク質、オステオネクチン、さまざまなリンタンパク質、石灰化プロセスに関与するプロテオリピド)、およびグリコサミノグリカンが見られます。 骨の基質には、骨の有機基質の原線維に関連して規則正しく配置されたヒドロキシアパタイトの結晶と、非晶質リン酸カルシウムが含まれています。 骨組織には 30 種類以上の微量元素 (銅、ストロンチウム、亜鉛、バリウム、マグネシウムなど) が検出され、体内の代謝プロセスで重要な役割を果たしています。 身体活動を体系的に増加させると、高度な石灰化により骨量が 10 ~ 50% 増加します。
骨組織は、高度に石灰化された細胞間物質を含む特殊なタイプの結合組織です。 骨格の骨はこれらの組織から構築されます。
細胞と細胞間物質の特徴。
骨組織は次のもので構成されています。
細胞:
1) 骨細胞 –分裂能力を失った主な数の骨組織細胞。 彼らは突起状の形態を持ち、細胞小器官が貧弱です。 にあります 骨の空洞、または ギャップ、骨細胞の輪郭に沿っています。 骨細胞のプロセスは骨細管に浸透し、その栄養性において役割を果たします。
2) 骨芽細胞 –骨組織を作る若い細胞。 骨では、それらは骨組織の形成と再生の場所である骨膜の深層に見られます。 これらの細胞はさまざまな形状 (立方体、錐体、または角張ったもの) をしており、1 つの核と、細胞質内によく発達した顆粒小胞体、ミトコンドリア、およびゴルジ複合体を含んでいます。
3) 破骨細胞 –石灰化した軟骨や骨を破壊する可能性のある細胞。 それらはサイズが大きく(直径は90ミクロンに達します)、3個から数十個の核を含みます。 . 細胞質はわずかに好塩基性で、ミトコンドリアとリソソームが豊富です。 顆粒小胞体は比較的発達が不十分です。
B. 細胞間物質、からなる:
主物質、比較的少量のコンドロイチン硫酸と、カルシウムと複合体(非晶質リン酸カルシウム、ヒドロキシアパタイト結晶)を形成するクエン酸やその他の酸が多く含まれています。
コラーゲン線維、小さな房を形成します。
細胞間物質、骨組織内のコラーゲン線維の位置に応じて、 機密扱いに:
1. 網状線維性骨組織。
2. 層状骨組織。 骨プレート
網状線維性骨組織。
その中でコラーゲン線維がランダムに配置されています。 この組織は主に胚に見られます。 成人では、頭蓋縫合部位や腱の骨への付着部位で発生します。
管状骨の骨幹を例にした層状骨組織の構造。
これは成人の体内で最も一般的なタイプの骨組織です。 で構成されています 骨プレート、骨細胞と特定の方向に配向したコラーゲン線維を持つ石化した非晶質物質によって形成されます。 隣接する層では通常、繊維の方向が異なるため、層状骨組織の強度が高まります。 骨格のほとんどの平らな骨や管状の骨の緻密で海綿状の物質は、この組織から作られています。
臓器としての骨。
骨は複数の組織からなる独立した器官であり、主なものは骨です。
管状骨の組織構造
それは骨端と骨幹で構成されます。 骨幹は外側から骨膜で覆われているか、 ストーマ膜(図6-3)。 骨膜には 2 つの層があります。 外側(線維性) – 主に線維性結合組織によって形成され、 内部(細胞) – 細胞が含まれています 骨芽細胞。骨に栄養を与える血管と神経は、骨膜とコラーゲン線維を通過します。 穴あき繊維。ほとんどの場合、これらの繊維は共通のプレートの外層でのみ分岐します。 骨膜は骨を周囲の組織と接続し、その栄養、発達、成長、再生に関与します。
骨幹を形成する緻密な物質は、特定の順序で配置された骨プレートで構成され、次の 3 つの層を形成します。
共通ラメラの外層。 彼の中で プレートは骨の骨幹の周囲に完全なリングを形成しません。 このレイヤーには以下が含まれます 穿孔チャネル、そこを通って血管が骨膜から骨に進入します。
平均,オステオン層 -血管の周りに同心円状に層になった骨プレートによって形成されます . このような構造はと呼ばれます オステオン、そしてそれらを形成するプレートは オステオンプレート。 オステオンは、管状骨の緻密な物質の構造単位です。 各オステオンは、いわゆるオステオンによって隣接するオステオンから区切られています。 胸の谷間ライン。骨骨の中心管には、結合組織を伴う血管が含まれています。 . すべての骨は通常、骨の長軸と平行に位置します。 骨管は互いに吻合します。 骨管にある血管は、骨髄および骨膜の血管と相互に連絡しています。 オステオンプレートに加えて、この層には次のものが含まれます。 インサートプレート(古い破壊されたオステオンの残骸) , オステオンの間にあります。
共通板の内層緻密な骨物質が髄腔に直接接する場所でのみよく発達します。
骨幹の緻密な物質の内部は、骨膜と同じ構造を有する骨内膜で覆われています。
米。 6-3. 管状骨の構造。 A. 骨膜。 B. 緻密な骨質。 B.エンドスト。 D. 骨髄腔。 1. 共通プレートの外層。 2. 骨層。 3. オステオン。 4. オステオンチャンネル。 5. プレートを挿入します。 6. 共通プレートの内層。 7. 海綿状組織の骨梁。 8. 骨膜の繊維層。 9. 骨膜の血管。 10. チャンネルを穿孔します。 11. 骨細胞。 (V. G. Eliseev、Yu. I. Afanasyevによるスキーム)。
トピックに関する記事