神経伝達物質とは何ですか? 主要な神経伝達物質 神経伝達物質の仕組み
神経伝達物質は、シナプスを介してニューロン(神経細胞)からニューロンにインパルスを伝達する内因性物質です。 神経伝達物質はシナプス小胞で生成され、シナプス間隙を通過した後、他のシナプスの受容体に取り込まれます。 神経伝達物質は、多くの単純な前駆体から合成されます。たとえば、十分な量が食物から得られ、少数の生合成プロセスを通じて吸収されます。 神経伝達物質は生命の鍵です。 その正確な数は不明ですが、100以上あることは間違いありません。
作用機序
神経伝達物質はシナプス小胞に存在し、シナプス小胞は軸索終末のシナプス前膜の下に存在します。 神経伝達物質はシナプス間隙を通じて生成および分布し、その後シナプス後膜の特定の受容体に結合します。 ほとんどの神経伝達物質のサイズはアミノ酸に匹敵しますが、一部の神経伝達物質はタンパク質やペプチドよりもさらに大きいものもあります。 神経伝達物質は、生成されてすぐに酵素によって代謝され、シナプス前ニューロンに取り込まれるか、シナプス後受容体に結合されます。 ただし、神経伝達を介してシナプス後反応を引き起こすには、通常、受容体への短期間の曝露で十分です。 活動電位または段階的な電位に応答して、シナプス前終末は神経伝達物質の産生を開始しますが、刺激がなくても少量の神経伝達物質が産生されます。 その後、神経伝達物質はシナプス後ニューロンの受容体に結合するまでシナプス中を移動します。 このプロセスにより、ニューロンが抑制されたり、興奮したりする可能性があります。 ニューロンは他のニューロンと関係を結ぶことができ、興奮性の効果が抑制性の効果を上回ると、それに応じてニューロンも興奮します。 その結果、軸索丘の新しい活動電位が現れ、神経伝達物質が放出され、隣接するニューロンへの情報伝達が刺激されます。
オープニング
20世紀初頭まで、科学者たちは脳内のほとんどのシナプス接続は電気的なものであると信じていました。 しかし、ラモン・イ・カハル(1852~1934)の組織学的研究中に、シナプス間隙として知られるニューロン間の距離が20~40nmであることが発見された。 このギャップの存在は、ニューロン間のコミュニケーションがそこを通過する化学伝達物質を介して行われることを示唆し、1921 年にドイツの薬学者オットー ローウィ (1873-1961) は、ニューロンが実際に特定の物質の生成を通じてコミュニケーションできることを確認しました。 カエルの脳神経を使った実験を通じて、ローウィ氏は神経周囲の生理食塩水の量を制限することでカエルの心拍数を下げることに成功した。 この実験の完了後、ローウィ博士は、特定の化学物質の濃度を変えることで心臓の機能を調節できると述べた。 さらに、オットー・ローウィは、最初に発見された神経伝達物質を発見しました。 ただし、一部のニューロンはギャップ結合を介した電気シナプスを介して通信し、特定のイオンが細胞間で直接移動できるようにします。
身元
神経伝達物質を決定するために、4 つの主な基準が開発されています。
物質はニューロン内で生成されるか、他の方法でニューロン内に侵入する必要があります。
ニューロンが活性化されると、物質が放出され、隣接するニューロンに特定の反応を引き起こす必要があります。
実験目的で、標的ニューロンに意図的に物質を注入した場合にも、同じ反応が起こるはずです。
作用機序は、物質を生成するニューロンから物質を除去することであるはずです。
薬理学、遺伝学、化学神経解剖学におけるすべての利点を考慮すると、「神経伝達物質」という用語は次のような物質に適用できます。
それらは、シナプス後膜を通過することによってニューロン間で信号を伝達します。
それらは膜張力にほとんど、またはまったく影響を与えず、たとえばシナプスの構造を変化させることによって単純な輸送機能も実行します。
それらは、伝達物質の生成と再吸収に影響を与える逆信号を送信することによって相互作用します。
神経伝達物質の解剖学的局在は、免疫細胞化学的アッセイを使用して決定できます。これにより、伝達物質または合成プロセスに関与する酵素のいずれかの位置がわかります。 さらに、このような分析を通じて、多くの伝達物質、特に神経ペプチドが局在していることを立証することができ、これは個々のニューロンがシナプス前終末から複数の伝達物質を生成する能力を示している。 染色、刺激、サンプリングなどのさまざまなアッセイや技術を使用して、中枢神経系の神経伝達物質を検出できます。
種類
神経伝達物質には多くの分類がありますが、最も便利なのはアミノ酸、ペプチド、モノアミンへの分類です。 主な神経伝達物質:
ペプチド: ソマトスタチン、サブスタンス P、コカインとアンフェタミンの正規化マトリックス、オピオイド ペプチド
ガス発生器: 一酸化窒素、一酸化炭素、硫化水素
さらに、50 を超える神経活性ペプチドが発見されており、このリストは常に更新されています。 それらの多くは、低分子量トランスミッターと一緒にリリースされます。 ただし、ペプチドがシナプス内の主要な伝達物質になる場合があります。 中枢神経系におけるオピオイド受容体との相互作用の特徴により、神経伝達物質ペプチドのかなりよく知られた例はβ-エンドルフィンです。 研究者の中には、一酸化窒素、一酸化炭素、硫化水素の分子などのガス分子だけでなく、個々のイオン (たとえば、シナプスで放出される) を神経伝達物質とみなしている人もいます。 ガスは神経細胞質で生成され、すぐに細胞膜を通って細胞間液および隣接する細胞に排出され、セカンドメッセンジャーの生成を刺激します。 溶解ガス神経伝達物質は非常に速く作用し、数秒しかかからずすぐに分解されるため、研究が困難です。 最も一般的な伝達物質はグルタミン酸で、人間の脳のシナプスを 90% 以上興奮させます。 次に来るのが GABA で、グルタミン酸を使用しないシナプスの 90% 以上を阻害します。 他の伝達物質はそれほど一般的ではありませんが、機能の点で非常に重要な場合があります。大部分の精神活性物質の効果は、一部の神経伝達物質システムの作用を変化させることによって発生します。 グルタミン酸や GABA 以外の伝達物質がこのプロセスに関与しています。 コカインやアンフェタミンなどの薬物はドーパミン系に大きな影響を与えます。 依存性薬物はオピオイドペプチドの機能的類似体として作用し、ドーパミンレベルを調節します。
行動
ニューロンは、神経インパルス (活動電位) が通過する神経ネットワークを形成します。 各ニューロンには、隣接するニューロンへの接続が 15,000 あります。 ただし、ニューロンは互いに接触しません(ギャップ結合を介した電気シナプスを考慮しない場合)。 代わりに、ニューロンは、神経伝達物質の助けを借りて神経細胞の隙間を通過するシナプスを介して相互に情報を伝達します。 実際、このプロセスは活動電位として知られる神経インパルスです。 シナプス前終末に到達すると、神経伝達物質の放出が刺激され、シナプス膜を通過してニューロンを興奮または抑制します。 新しいニューロンはそれぞれ他の多くのニューロンと接続されており、興奮性効果の合計が抑制性効果を上回る場合、それに応じてニューロンは興奮します。 これにより、軸性丘の新しい活動電位が生成され、ニューロンからニューロンへ情報を伝達する神経伝達物質が放出されることは注目に値します。
興奮作用と抑制作用
神経伝達物質は、さまざまな形でニューロンの機能に影響を与える可能性があります。 それにもかかわらず、ニューロンの電気的興奮性に影響を与えることができるのは、興奮または抑制の 2 つの方法のみです。 神経伝達物質は膜を通過するイオンの流れを調節し、それによって活動電位を生成する細胞の能力を増加 (興奮) または減少 (抑制) します。 したがって、シナプスには非常に多様な種類があるにもかかわらず、それらはすべてこれら 2 つの状態に関する情報のみを伝え、対応する名前を持っています。 最初のタイプのシナプスは興奮性ですが、2 番目のタイプのシナプスは抑制性です。 それらは外部的には互いに異なり、影響を受けたニューロンの異なる部分に位置します。 ニューロンは毎秒、何千もの興奮性信号と抑制性信号を同時に受け取ります。 最初のタイプの丸いシナプスは通常、樹状突起の内側に位置し、2 番目のタイプの平らなシナプスは細胞の外側に位置します。 さらに、最初のタイプのシナプスは、より緻密な構造とより広いシナプス ギャップを持っています。 そして最後に、それらの活性ゾーンも 2 番目のタイプのシナプスの活性ゾーンよりも大きくなります。 それらの別々の配置により、ニューロンは 2 つの部分、つまり興奮性樹状突起ツリーと抑制性細胞体に分割されます。 抑制に関して言えば、興奮は樹状突起から発生し、軸索丘に伝播し、それによって活動電位が発火します。 このメッセージを止めるには、活動電位の発生部位である丘にできるだけ近い細胞体を抑制するのが最善です。 言い換えれば、抑制は興奮活性化の瞬間を決定することにあります。 通常の状態では、細胞体は抑制されており、軸索丘で活動電位を生成する唯一の方法は抑制を停止することです。 これは比喩的に次のように説明できます。興奮信号は競走馬のようなもので、いつでも解き放たれる準備ができていますが、そのためには抑制の門が開く必要があります。
神経伝達物質の影響の例
上で述べたように、神経伝達物質の唯一の直接的な目的は受容体を活性化することです。 したがって、神経伝達の影響は、このプロセスに関与するニューロンの接続と、伝達物質が結合する受容体の化学的性質に依存します。 神経伝達物質の重要な影響の例をいくつか示します。
-
薬物使用の影響
神経伝達物質に対する薬物の影響を理解することは、神経科学分野の研究に大きく依存します。 ほとんどの神経科学者は、そのような研究が多くの神経疾患や障害の原因を理解し、それらと戦う効果的な方法を見つけ、さらにはそれらを予防または完全に治療する方法を見つけるのに役立つと信じています。 薬は患者の行動や神経伝達物質の活動の変化に影響を与える可能性があります。 たとえば、その組成に含まれる合成酵素は、神経伝達物質の合成を減少させたり、完全にブロックしたりする可能性があります。 これが起こると、活性な神経伝達物質の数が劇的に減少します。 一部の薬物は、特定の種類の神経伝達物質の産生をブロックまたは刺激することができますが、他の薬物はシナプス小胞への神経伝達物質の蓄積を妨げ、膜での神経伝達物質の保持を不可能にします。 神経伝達物質がその受容体に結合するのを妨げる薬剤は、受容体拮抗薬と呼ばれます。 たとえば、クロルプロマジンなどの薬物は、脳内のドーパミン受容体拮抗薬です。 受容体アゴニストとして知られる他の薬物の成分は、受容体自体に結合し、実際の神経伝達物質を模倣します。 そのような薬物の例はベンゾジアゼピンであり、これは作用を模倣し、それによって患者の不安を軽減します。 他の薬物は、神経伝達物質が活性化された後にそれを不活性化し、それによって持続時間を延長します。 これは、再取り込みを防ぐか、破壊酵素を阻害することによって達成できます。 最後に、薬物は中枢神経系および末梢神経系のニューロン活動をブロックすることによって活動電位を防ぐこともできます。 テトロドトキシンなどの神経活動を遮断する薬物の使用は、多くの場合致命的です。 主要なシステムの神経伝達物質を標的とする薬剤はシステム全体に影響を与えるため、その作用が複雑になります。 たとえば、コカインはシナプス前ニューロンによるドーパミンの再取り込みをブロックし、神経伝達物質がシナプス間隙に長時間留まるようにします。 ドーパミンが必要以上に長くシナプス内に存在するため、神経伝達物質はシナプス後ニューロンの受容体に結合し続け、心地よい感情状態を引き起こします。 コカインへの身体的中毒は、シナプスでのドーパミンの長期放出が原因で、特定のシナプス後受容体の数が減少します。 物質の効果が終わると、神経伝達物質と受容体との相互作用が減少するため、患者はうつ病になります。 選択的セロトニン再取り込み阻害剤 (SSRI) であり、実際、シナプス前細胞によるセロトニンの再取り込みをブロックし、その結果、シナプス内のセロトニンの量が増加し、その結果、物質がシナプスに残ります。必要以上に長く使用すると、体自体によるセロトニンの生成が増加します。 アルファ-メチル-P-チロシン (AMPT) は、チロシンがドーパミン前駆体である L-デヒドロキシフェニルアラニンに変換されるのを防ぎます。 レセルピンは小胞内でのドーパミンの蓄積を防ぎ、デプレニルはモノアミンオキシダーゼ-Iを阻害することでドーパミンレベルを上昇させます。
アゴニスト
アゴニストは、神経伝達物質を含む受容体に結合し、それによって内部物質の結合と同じ反応を引き起こすことができる化学物質です。 神経伝達物質アゴニストは、伝達物質と同じ受容体応答を引き起こします。 筋肉がリラックスした状態のときに機能します。 アゴニストには、直接アゴニストと間接アゴニストの 2 種類があります。
直接作用性アゴニストは、受容体の活性部位に直接結合することにより、神経伝達物質のように作用します。 これにより、投与を受けた人は、あたかも脳に直接注射されたかのように薬の効果を体験することができます。 これらには、アポモルヒネおよび。
間接アゴニストは、神経伝達物質の産生を刺激することにより、神経伝達物質の作用を強化します。 その一例がコカインです。
薬物アゴニスト
「アゴニストは、(受容体の活性部位に結合することによって)受容体に作用し、特定の生物学的反応を引き起こす化合物または内部物質です(独自の内部活性を持っています)。 化学アゴニストの受容体への結合は、内部物質 (ホルモンや神経伝達物質など) が同じ受容体に結合することによって生じる心理的反応と同様の心理的反応を模倣します。 多くの場合、生物学的反応は相互作用可能なアゴニストの濃度に依存します。 濃度が増加すると、結合する受容体の数も増加し、それに応じて生物学的反応も増加します。 生理学的反応の強さは、注入された薬物の量と受容体の結合の強さに直接依存します。 ほとんどの薬物は複数の受容体と相互作用し、相互作用します。」 タバコに含まれるアセチルコリンニコチン受容体作動薬です。 オピオイドアゴニストは、ヘロイン、ヒドロコドン、オキシコドン、コデイン、およびメタドンです。 これらの薬剤は、通常、エンケファリンなどの内部伝達物質にのみ反応するミューオピオイド受容体を活性化します。 このような受容体が活性化されると、人は多幸感、痛みの軽減、眠気を経験します。
アンタゴニスト
アンタゴニストは、体内で作用して別の化合物(アヘン剤など)、特に神経系を抑制し自然に生成される化合物の生理活性を低下させる化合物です。 アンタゴニストの作用機序は、神経受容体に結合してブロックすることです。 このメカニズムは筋肉が収縮するときに働きます。 アンタゴニストには、直接アンタゴニストと間接アンタゴニストの 2 種類があります。
直接作用するアンタゴニストは神経伝達物質ではなく受容体と相互作用し、その結果、受容体に結合する能力を失います。 最も有名な敵対者は。
間接的アンタゴニストは、神経伝達物質の放出/産生を阻害します。 一例はレセルピンです。
薬物拮抗薬
薬物アンタゴニストは受容体に結合し、受容体に特定の生物学的反応を引き起こします。 したがって、薬の拮抗薬はそれ自体の活性を持たないと彼らは言います。 アンタゴニストは、アゴニスト(薬物、ホルモン、神経伝達物質など)が受容体に結合するのを妨げることによってその作用をブロックするため、受容体「ブロッカー」とも呼ばれます。 アンタゴニストは競合性と不可逆性に分けられます。 競合的アンタゴニストは、受容体への結合に関してアゴニストと競合します。 アンタゴニストの濃度が増加すると、アゴニストの可能性が減少し、生理学的反応が低下します。 高濃度のアンタゴニストは、この反応を完全に阻害することさえできます。 ただし、アゴニストの濃度を高めるだけで阻害を元に戻すことができます。 競合アンタゴニストの存在下では、競合物質が存在しない場合と同じ反応を引き起こすためには、はるかに高い濃度のアゴニストが必要となります。 不可逆的なアンタゴニストは受容体に非常に強く結合しているため、アゴニストはそれと戦うことができません。 このようなアンタゴニストは、受容体と共有化学結合を形成することさえ可能です。 いずれにせよ、十分な濃度の不可逆的なアンタゴニストを使用すると、結合していない残りの受容体の数が非常に少なくなり、どの濃度のアゴニストでも最大の生物学的反応を引き出すことができなくなります。
前駆体
神経伝達物質前駆体の取り込みによって神経伝達物質の合成が増加することは確かですが、その過程で神経伝達物質の産生が増加するかどうか、またシナプス後受容体の興奮性が増加するかどうかはまだ確立されていません。 神経系は神経伝達物質合成の増加などの変化に適応して永続的な興奮状態をもたらす可能性があるため、産生が増加したとしても、これが神経伝達物質シグナルの強度に影響を与えるかどうかは不明です。 いくつかの神経伝達物質がうつ病に関与しており、その前駆体がうつ病と戦うのに効果がある可能性があるという証拠があります。
カテコールアミンおよび微量アミンの前駆体
L-デヒドロキシフェニルアラニンは、血液脳関門を通過できるドーパミン前駆体であり、パーキンソン病の治療に使用されます。 しかし、神経伝達物質前駆体の投与は、うつ病やノルアドレナリン濃度の低い患者にはあまり効果がありません。 L-フェニルアラニンとL-チロシンはドーパミン、ノルエピネフリン、エピネフリンの前駆体であり、ビタミンB6、ビタミンC、S-アデノシルメチオニンに依存しています。 いくつかの研究によると、L-フェニルアラニンとL-チロシンは抗うつ薬である可能性がありますが、正確な確証はまだ見つかっていません。
セロトニン前駆体
病気と障害
病気や障害も神経伝達物質システムに影響を与える可能性があります。 たとえば、ドーパミン生成の中断はパーキンソン病を引き起こす可能性があり、パーキンソン病は人に不随意な動きを引き起こし、しびれ、震え、震え麻痺、その他の症状を引き起こします。 いくつかの研究によると、ドーパミンレベルが低すぎると統合失調症や、 さらに、うつ病患者ではセロトニンレベルも低下します。 最も一般的なものは、ニューロンによるセロトニンの処理または取り込みをブロックし、その結果、より多くのセロトニンがシナプスに残り、最終的に患者の気分を正常化します。 さらに、グルタミン酸の生成または吸収の障害は、次のような多くの精神障害を引き起こす可能性があります。
グルタミン酸は、脳や脊椎で機能するさまざまな興奮性シナプスに関与しています。 また、多くの「プラスチック」シナプスの一部でもあります。 増えたり減ったりするもの。 プラスチックのシナプスが記憶の主な保管場所であると考えられています。 グルタミン酸が過剰に生成されると脳が過剰に興奮し、興奮毒性や細胞死につながり、ひいては発作や脳卒中を引き起こす可能性があります。 興奮毒性は、虚血性脳卒中、てんかん、ハンチントン舞踏病などのいくつかの慢性疾患を引き起こす可能性があります。
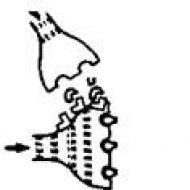
シナプスでは、神経インパルスの伝達プロセスは、シナプス小胞に蓄積する神経伝達物質(神経ホルモン)の助けを借りて起こります。神経伝達物質は、シナプス間隙へのニューロン伝達中に放出され、シナプス後膜の特定の受容体(つまり、シナプス小胞)に付着します。 「錠の鍵のようにフィットする」領域)。」)。 シナプス後膜の透過性の変化の結果、信号はあるニューロンから別のニューロンに伝達されます。 メディエーターは、シナプスのレベルで神経信号の伝達をブロックし、シナプス後ニューロンの興奮性を低下させる可能性があります。 神経伝達物質の不活性化は、発酵 (酵素による破壊) とシナプス前末端への逆吸収の 2 つの方法で起こります。 これにより、次のパルスが到着するまでにバブル内の在庫が回復します。
1 - 神経インパルス、2 - X 物質の分子、3 - 受容体部位、4 - 神経伝達物質分子
神経伝達物質分子はニューロン I の末端プラークから放出され、ニューロン II の樹状突起上の特定の受容体に結合します。 X 物質の分子の構成はこれらの受容体に適合せず、シナプス効果を引き起こしません。
シナプスの興奮性または抑制性の機能は、シナプスによって分泌されるメディエーターの種類と、シナプス後膜に対するメディエーターの作用に依存します。 いくつかの神経伝達物質は興奮作用のみを持ち、他の神経伝達物質は抑制(抑制)作用のみを持ちますが、他の神経伝達物質は神経系の一部で活性化因子の役割を果たし、他の部分では抑制因子の役割を果たします。
神経伝達物質の機能。現在、数十種類の神経伝達物質が知られていますが、その機能はまだ十分に研究されていません。
アセチルコリン
すべての神経伝達物質の中で、アセチルコリンは最初に発見されたものの 1 つです。 これはニューロンと筋細胞の接合部に存在し、筋収縮に関与し、心臓と呼吸リズムの低下を引き起こします。 それは酵素アセチルコリンエステラーゼによって不活化されます。 アセチルコリンは脳の活動において重要な役割を果たしていますが、他のほとんどの神経伝達物質と同様に、その機能は完全には理解されていません。 喉の渇きの感覚の重要な調節因子であることが知られています。 おそらく、アセチルコリンも記憶システムの重要な要素であると考えられます。 アルツハイマー病は、間脳核内のアセチルコリンおよびコリン作動性受容体の機能障害に関連しています。
モノアミン
モノアミンは、同じアミノ基の一部であるノルアドレナリン(ノルエピネフリン)、ドーパミン、セロトニンの 3 つの重要な神経伝達物質と呼ばれます。
ノルアドレナリン
大脳皮質の覚醒を担当し、感情の高揚、空腹、心拍数の増加に伴う身体的変化を調節します。 恐怖に発展する不安の感情状態は、ノルアドレナリン交換の違反と関連しています。
セロトニン
脳のあらゆる部分に存在し、睡眠の調節に重要な役割を果たし、感覚経路を循環する情報量を決定します。 憂鬱な状態はセロトニン代謝の違反と関連しています。
ドーパミン
選択的注意のプロセス、身体部分の調整された動きに参加し、大脳辺縁系の「快楽中枢」と網様体形成のいくつかの核に存在します。 被殻および縫線核(基底核)におけるドーパミンの欠乏は、パーキンソン病の主な原因である可能性があります。 ドーパミン代謝の違反は、統合失調症発症の生化学的根拠です。 コカインやアンフェタミンなどの興奮剤は、脳内のドーパミン作動性活動を増加させます。
これらの機能に加えて、モノアミンは気分や感情障害と密接に関連しています。 臨床的うつ病は、モノアミン、特にノルアドレナリンとセロトニンのレベルの変化によって発生します。
モノアミンの部分的な不活化は、モノアミンオキシダーゼ酵素による酸化の結果として起こります。 このプロセスにより、脳の活動が正常なレベルに戻ります。
ガンマアミノ酪酸 (GABA)
抑制性神経伝達物質。 その作用は主に、神経インパルスに関連した脳ニューロンの興奮性を低下させることにあります。 GABA (GABA) と同様に、バルビツール酸塩、精神安定剤、アルコールなどの古典的な抑制剤が作用します。
エンドルフィン
1975年、内因性オピオイドペプチド(エンドルフィン、ダイノルフィン、エンケファリン)、つまり「脳自体のモルヒネ」が発見されました。 体内でのそれらの機能は多様であり、まだ完全には理解されていませんが、これらの物質が痛みを和らげるのに役立つことは間違いありません。 これらは、痛みの知覚を阻害する複雑なシステムの神経伝達物質です。 これらは特定のオピオイド受容体 (5 種類) と相互作用し、体外から投与されたオピオイドも同様に反応します。 オピオイドのメカニズムに関する既存の考えでは、オピオイドに対する耐性と依存の発生を説明することはまだできません。

神経伝達物質とともに、次のグループがあります。 神経調節物質神経反応の調節に関与し、メディエーターと相互作用してその効果を調節します。 例には、痛みのシグナル伝達に関与するサブスタンス P やブラジキニンが含まれます。 しかし、脊髄のシナプスでのこれらの物質の放出は、エンドルフィンとエンケファリンの分泌によって抑制されるため、痛みの神経インパルスの流れが減少します。
神経調節物質は軸索の末端に作用し、神経伝達物質の放出を促進または阻害します。
神経調節物質の機能は、睡眠プロセスにおいて重要な役割を果たす因子 8 などの物質によって実行されます。 コレシストキニン、満腹感を司る。 喉の渇きを調節するアンジオテンシンなど。
最も有名な 3 つの神経伝達物質。これなしでは私たちの生活はただうんざりするでしょう。
神経伝達物質は常にあなたとともにある休日です。それらが喜びや喜びの感情を与えるということはよく耳にしますが、それらがどのように作用するかについてはほとんどわかっていません。
私たちは 3 つの最も有名な神経伝達物質について話します。これらの神経伝達物質がなければ、私たちの生活は単にうんざりするでしょう。
神経伝達物質の仕組み
神経細胞は、軸索と樹状突起というプロセスの助けを借りて相互に通信します。 それらの間のギャップは、いわゆるシナプス間隙です。 ここでニューロンが相互作用します。
メディエーターは細胞内で合成され、軸索の端、つまりシナプス前膜に送られます。 そこで、電気インパルスの作用下で、それらはシナプス間隙に入り、次のニューロンの受容体を活性化します。 受容体の活性化後、神経伝達物質は細胞に戻るか(いわゆる再取り込みが起こります)、または破壊されます。
神経伝達物質自体はタンパク質ではないため、「ドーパミン遺伝子」や「アドレナリン遺伝子」は存在しません。タンパク質はすべてのサポート作業を行います。
- 酵素タンパク質は神経伝達物質の物質を合成し、
- トランスポータータンパク質は配送を担当し、
- 受容体タンパク質は神経細胞を活性化します。
1 つの神経伝達物質が正しく機能するためには、複数のタンパク質が関与している可能性があります。これは、複数の異なる遺伝子を意味します。
ドーパミン
ドーパミンは、脳のさまざまな領域のニューロンを活性化することにより、いくつかの役割を果たします。
- まず、運動活動を担当し、動く喜びを与えます。
- 第二に、新しいことを学ぶとほとんど子供じみた喜びを感じ、新しいものを探したいという欲求を与えます。
- 三番目、ドーパミンは、報酬とモチベーションの強化という重要な機能を果たします。私たちが人類の生命にとって有益なことを行うとすぐに、ニューロンは私たちに報酬、つまり満足感(快感とも呼ばれます)を与えます。
基本的なレベルでは、私たちは人間の単純な喜び、つまり食事やセックスに対して報酬を受け取りますが、一般に、満足を達成するためのオプションは全員の好みに依存します。完成したコードに対して誰かが「ニンジン」を得るでしょうし、誰かがこれに対して「ニンジン」を得るでしょう。記事。
報酬システムはトレーニングに関連しています。人は喜びを感じ、脳内に新たな因果関係が形成されます。 そして、その喜びが過ぎ去り、どうすれば再びそれを得ることができるかという疑問が生じたとき、別の記事を書くという簡単な解決策があります。
ドーパミンは、仕事や勉強にとって素晴らしい興奮剤であると同時に、理想的な薬物のように見えます。ほとんどの薬物(アンフェタミン、コカイン)はドーパミンの作用に関連していますが、深刻な副作用があるだけです。
- ドーパミンの「過剰摂取」リード 統合失調症に(脳は非常に活発に機能するため、幻聴や幻視が現れ始めます)、
- 欠陥- うつ病性障害またはパーキンソン病の発症 .
ドーパミンには、D1 から D5 までの番号が付けられた 5 つの受容体があります。 4 番目の受容体は、新規性の探索を担当します。 これは DRD4 遺伝子によってコードされており、その長さがドーパミン取り込みの強度を決定します。
繰り返しが少ないほど人は快楽の頂点に達しやすくなります。 そういう人は美味しい夕食と良い映画を観れば十分でしょう。
繰り返しが多ければ多いほど- 最大 10 個まで存在する可能性があります - 楽しむのは難しくなります。 そのような人々は、報酬を得るために一生懸命働かなければなりません。たとえば、世界一周旅行に行く、山の頂上を征服する、バイクで宙返りする、ラスベガスで全財産を賭けるなどです。 このような遺伝子型は、古代人のアフリカからユーラシア全域への移動範囲に関連しています。
悲しい統計もあります。 DRD4 の「不十分な」亜種は、重大な犯罪で刑務所で有罪判決を受けた人々の間でより一般的です。
ノルアドレナリン
ノルアドレナリンは、覚醒と素早い意思決定のための神経伝達物質です。ストレスがかかると活性化し、極限状況では「闘争・逃走」反応に関与します。
ノルアドレナリンはエネルギーの急増を引き起こし、恐怖感を軽減し、攻撃性のレベルを高めます。
体細胞レベルでは、ノルアドレナリンの影響により、心拍数が速くなり、血圧が上昇します。
ノルアドレナリンは、サーファー、スノーボーダー、モーターサイクリスト、その他のエクストリーム スポーツ愛好家だけでなく、カジノやゲーム クラブの愛好家にも好まれている仲介者です。脳は現実の出来事と想像上の出来事を区別できないため、財産を失うという命の危険はありません。ノルアドレナリンを活性化するには、カードに含まれるだけで十分です。
- 上級ノルアドレナリンは視力と分析能力の低下を引き起こし、
- 欠陥- 退屈と無関心へ。
SLC6A2 遺伝子は、ノルエピネフリン トランスポーター タンパク質をコードします。 これは、ノルエピネフリンのシナプス前膜への再取り込みを確実にします。 危険な状況にうまく対処した後、ノルアドレナリンが人体内でどのくらいの時間作用するかは、その働きによって決まります。 この遺伝子の変異は、注意欠陥障害 (ADHD) を引き起こす可能性があります。
セロトニン
私たちはそれを「幸福のホルモン」として聞き慣れていますが、 セロトニンはホルモンではない、そして「幸福」に関してはすべてはそれほど単純ではない.
セロトニンは、ポジティブな感情をもたらすだけでなく、ネガティブな感情への感受性を低下させる神経伝達物質です。 「隣接する」神経伝達物質であるノルエピネフリンとドーパミンをサポートします。
セロトニンは運動活動に関与し、全体的な痛みを軽減し、体の炎症との闘いを助けます。
また、セロトニンは脳内の活動信号の伝達の精度を高め、集中力を高めます。
- セロトニンが多すぎる(たとえば、LSD を使用している場合) 脳内の二次信号の「音量」が増加し、幻覚が発生します。
- セロトニン不足そしてポジティブな感情とネガティブな感情の間の不均衡がうつ病の主な原因です。
5-HTTLPR 遺伝子は、セロトニントランスポータータンパク質をコードします。 遺伝子配列には反復領域が含まれており、その数は異なる場合があります。
- チェーンが長ければ長いほど人はポジティブな姿勢を維持し、ネガティブな感情から切り替えることが容易になります。
- 短いほど- 否定的な経験がトラウマになる可能性が高くなります。
繰り返しの回数も関係する 乳児突然死症候群、アルツハイマー病の発症における攻撃的な行動、およびうつ病の傾向.

神経伝達物質の破壊
神経伝達物質の作用は、休日のようなもので、まるで花火を見にみんなで楽しい群衆の中で通りに繰り出すようなものです。 しかし、この休日は永遠に続くことはできませんし、そうすべきではありません。夜空のネオンローズは、見慣れた星座や夜明けに取って代わられるに違いありません。
これを行うために、体はメディエーターを再取り込む機能を持っています。このとき、物質がシナプス間隙から軸索のシナプス前膜に戻り、神経伝達物質の作用が停止します。
しかし、再摂取だけでは不十分な場合があり、より効果的な手段、つまり神経伝達物質分子の破壊が必要になることがあります。
これらの機能もタンパク質によって実行されます。
COMT 遺伝子は、ノルエピネフリンとドーパミンを破壊する酵素カテコール-O-メチルトランスフェラーゼをコードしています。 タンパク質の働きは、ストレスの多い状況にどれだけうまく対処できるかによって決まります。
- 活性型COMT遺伝子の所有者- 本質的に戦士 - 情報処理と快感を司る脳の前頭葉で受け取るドーパミンのレベルが低下します。 そのような人々は、ストレスの多い状況によりよく適応し、コミュニケーションにオープンで、より良い記憶力を持っています。 しかし、ドーパミンレベルが低いため、人生の楽しみが減り、うつ病になりやすく、運動機能もあまり発達していません。
- COMT 遺伝子の不活性変異体状況を逆転させます。 不活性な突然変異を持つ人は、優れた運動能力を持ち、より創造的ですが、痛みにあまり耐えられず、ストレスの多い状況に陥るとすぐに、過敏性、衝動性、不安に陥ります。
COMT遺伝子の変異も関連している パースキンソン症と高血圧を伴う.
 モノアミンオキシダーゼA酵素遺伝子MAOAモノアミン - アドレナリン、ノルエピネフリン、セロトニン、メラトニン、ヒスタミン、ドーパミンなどの 1 つのアミノ基を持つ神経伝達物質の不活性化に関与します。 MAOA 遺伝子の働きが良くなればなるほど、ストレスの多い状況によって引き起こされる心の霧がより早く中和され、人は情報に基づいた意思決定をより早く下せるようになります。
モノアミンオキシダーゼA酵素遺伝子MAOAモノアミン - アドレナリン、ノルエピネフリン、セロトニン、メラトニン、ヒスタミン、ドーパミンなどの 1 つのアミノ基を持つ神経伝達物質の不活性化に関与します。 MAOA 遺伝子の働きが良くなればなるほど、ストレスの多い状況によって引き起こされる心の霧がより早く中和され、人は情報に基づいた意思決定をより早く下せるようになります。
MAOA 遺伝子と呼ばれることもあります。 「犯罪者のゲノム」 : 特定の遺伝子変異が病的攻撃性の発生に寄与します。 この遺伝子は X 染色体上にあり、女の子はこの遺伝子のコピーを 2 つ持っているのに対し、男の子は 1 つしか持っていないため、統計的には男性の方が「生まれながらの犯罪者」が多いのです。
すべてを遺伝のせいにするのはやめましょう- 「猛烈な」MAOA 遺伝子に関してさえ、すべてが簡単ではありません。ニュージーランドの科学者による研究では、この遺伝子と攻撃的行動との関連性は、トラウマ的な経験が存在する場合にのみ現れることが示されました。
神経伝達物質がどのように機能するかの原理を理解することで、習慣的な感情や気分の変化を新たに見つめることができ、実際に何が私たちの性格を形作るのかについての考えを再考することさえ可能になります。出版された
喜び、悲しみ、恐怖、疑い、喜び、これらすべてを私たちは人生の特定の瞬間に感じることができます。 しかし、これらの感情はどこから来るのでしょうか? 昔は、すべての始まりは心臓か胃だと誰もが信じていました。
しかし、人々がもう少し賢くなると、さまざまな感情を引き起こす化学プロセスが脳内で起こっていることに気づきました。 そして体は心臓に従うのではなく、中枢神経系と自律神経系に従うのです。
人間の脳には膨大な数のニューロンがあるという事実は、人々が比較的昔から認識していました。 しかし、60年代になって初めて、神経生物学者は神経伝達物質という非常に重要な発見をしました。
神経伝達物質とは何ですか?またその目的は何ですか?
脳細胞ニューロンには、神経インパルスを伝達することによって互いに相互作用する多くのプロセスがあります。 これらのシナプス接続は非常に多数です (平均して 1 細胞あたり約 10,000 個のプロセス)。
さらに、ニューロンは密接に接続されていません。 プロセス間には小さなシナプスギャップがあり、神経インパルスが放電の形で通過します。 しかし、後に、複雑な化学反応を起こすための単純な衝動だけでは十分ではないことが判明しました。

ここで神経伝達物質が活躍します。 それらはニューロンのまさに接合部、つまりシナプス接続で形成されます。 神経伝達物質は、ニューロンからのインパルスを筋肉系全体に伝えます。 そして、それぞれに独自の特性と機能があります。
あなたが悲しいとき、または明るい気分のとき、これは調停者の仕事です。 神経細胞を元気にしたり穏やかにしたりするのは彼らです。
現在までに、膨大な数の神経伝達物質が特定されています。 しかし、それらの多くはまだ調査の余地があります。 この記事では、主な神経伝達物質と、それらが私たちの体に及ぼす影響について説明します。
主な神経伝達物質:その機能と特徴。

グルタミン酸
グルタミン酸それはアミノ酸であり、神経系の主要な興奮性神経伝達物質です。 そのため、私たちの脳は、まるで一度に数杯のコーヒーを飲んだかのように、興奮した状態で働きます。 この神経伝達物質は、新しい情報の獲得と同化に貢献します。
過剰なグルタミン酸は悪影響を引き起こす可能性があります。 突然の発作の後、グルタミン酸の急増により直接的な後遺症が残ります。
ギャバ
ギャバ- アミノ酸、中枢神経系の主な鎮静神経伝達物質です。 ガンマアミノ酪酸は、グルタミン酸レベルが上昇している不安な状況(たとえば、難しい面接や試験の前)で心を落ち着かせるのに役立ちます。 また、新陳代謝を調節し、睡眠の質を向上させます。 GABA 欠乏症の症状については、記事 4 自分で特定できる GABA 欠乏症の症状をご覧ください。
神経伝達物質の発見の背後には、ドイツ人の O. レヴィ、ロシアの A.F. サモイロフ、イギリス人の G. デールの研究がありました。 信じられないことに、O. レヴィは、夢の中で神経伝達物質の存在を証明するのに役立つ実験の計画を見ました。 1936 年、科学者たちはこの発見によりノーベル賞を受賞しました。
アドレナリン
アドレナリン- 脈拍を速め、血圧を上げ、呼吸を速め、腸の収縮を減らすホルモン。 ストレスの多い状況で放出されると、体力と持久力が高まりますが、同時に知的能力が一時的に鈍くなります。 多くの初心者登山家やスカイダイバーが、緊急事態の瞬間を思い出して、すべてが非常に早く起こり、「霧の中のようだった」と言うのは、アドレナリンのせいです。
ノルアドレナリン
ノルアドレナリンアドレナリンに似ていますが、より楽しいものです。 たとえば、事故の後(アドレナリンが爆発したとき)、私たちは気分が悪くなり、そのような状況にはもう陥りたくなくなります。
しかし、高い山をスキーで滑り降りたり、崖の上に立ったり、騒がしいディスコで踊ったりするときにも、私たちはストレスを感じます。 私たちはそれが大好きで、もっと欲しいと思っています。 ノルアドレナリンは喜びと不安の組み合わせです。

ドーパミン
ドーパミンノルアドレナリンに先行して、ほぼ同じように体に影響を与えます。 ただし、この神経伝達物質はモチベーションが高まったときに発生します。 たとえば、明日はずっと欲しかった車を買うだろう、給料日が来る、待ち望んでいた休暇が来る、などと長い間考えているとき。 ドーパミンのせいで、寝る前に煩わしい考えに悩まされて目が覚めてしまうことがあります。
セロトニン
セロトニン- ほぼ最も愛され有名なホルモン。 胃腸管の機能を調節し、筋肉の緊張を維持し、身体活動を促進します。 しかし、重要なことは、彼のおかげで私たちはいつも良い気分です。
セロトニンのレベルが低いと、私たちは憂鬱になり、情緒が不安定になります。 体はグルコースとトリプトファンからこのホルモンを合成します。 これらの物質は、お菓子、果物、チョコレートから簡単に摂取できます。 おそらく、ストレス時に私たちがこれらの食品に頼ってしまうのは、セロトニンが不足しているためかもしれません。
メラトニン
メラトニン- 概日リズムに関与するホルモン。 光にさらされると、この神経伝達物質の合成が減少します。 メラトニン合成レベルは通常20時から増加し始め、3時にピークに達します。 このホルモンは、一晩中ぐっすりと快適に眠るのに役立ちます。 そのため、21時以降は光量を減らして23時までに就寝するようにしましょう。
メラトニンは通常、25~30歳で合成されます。 さらにその生成量が減少し、老化につながります。 メラトニンはホルモン系にも影響を与え、性的活動を増加させ、老化を遅らせ、月経周期、血圧、消化、脳細胞を調節します。

エンドルフィン
エンドルフィン。身体のための天然薬である快楽ホルモンのセット全体のよく知られた名前。 エンドルフィンはさまざまなプロセスに関与しており、快感や多幸感を引き起こし、情報の記憶を助け、空腹感を調節します。
さらに、エンドルフィンは痛みを軽減するのに役立ちます。 アメリカの研究で実験が行われました。 妊婦は出産の1~2週間前に好きな音楽を直接聞くことが許可された。 その結果、彼らの多くは出産時の痛みを大幅に軽減し、鎮痛剤をまったく拒否した人もいます。
ご覧のとおり、神経伝達物質は私たちの生活の中で驚くべき役割を果たしています。 それらは私たちの現実認識に直接影響を与え、体内の一連の重要なプロセスを調節します。
自律神経の仕組みを知れば、自分の気分をコントロールできるようになります。 そうすることで、うつ病を回避し、新たな高みに到達するためのモチベーションを高め、常に良い気分を保つことができます。
神経伝達物質 (神経伝達物質、メディエーター、英語のメディエーター - メディエーターから)- 電気インパルスが神経細胞からニューロン間のシナプス空間(ギャップ)を通って伝達され、また、例えばニューロンから筋肉組織に伝達される、低濃度で高い生理活性を有する物質。 シナプス前終末に入る神経インパルスにより、メディエーターがシナプス間隙に放出されます。 メディエーター分子は細胞膜の特定の受容体タンパク質と反応し、膜貫通イオン流の変化を引き起こす一連の生化学反応を開始し、膜脱分極と活動電位の出現につながります。
ニューロンは互いに電気インパルスを伝達しますが、ニューロンの間には誘電体である空間があり、メディエーターは別のニューロンに信号を伝達するためにこの空間を通過する必要があります。
この設計により、複雑な信号 (コンピューターの場合とは異なり、Yes/No のみではなく、メディエーターの約 24 の組み合わせ) を送信することができます。これらの信号は、私たちが知覚する現実全体を組み合わせの接続で送信します。 メディエーターはニューロン間の仲介者であり、記憶、感覚、知覚を保存する役割を果たします。
伝統的に、神経伝達物質はアミノ酸、ペプチド、モノアミン(カテコールアミンを含む)の 3 つのグループに分類されています。
アミノ酸
- GABA は、ヒトおよび哺乳類の中枢神経系における最も重要な抑制性神経伝達物質です。
- グリシン - 神経伝達物質のアミノ酸として、二重の効果があります。 グリシン受容体は脳と脊髄の多くの領域に存在します。 グリシンは受容体に結合することにより、ニューロンに「抑制」効果を引き起こし、ニューロンからのグルタミン酸などの「興奮性」アミノ酸の放出を減少させ、GABA の放出を増加させます。 グリシンはまた、NMDA 受容体の特定の部位に結合するため、興奮性神経伝達物質のグルタミン酸およびアスパラギン酸からのシグナル伝達を促進します。 脊髄では、グリシンは運動ニューロンの抑制につながり、神経内科診療でグリシンを使用して筋緊張の増加を解消することができます。
- グルタミン酸 (グルタミン酸) は、脊椎動物の神経系、小脳および脊髄のニューロンにおいて最も一般的な興奮性神経伝達物質です。
- アスパラギン酸(アスパラギン酸)は、大脳皮質のニューロンにおける興奮性神経伝達物質です。
カテコールアミン
- アドレナリンは興奮性神経伝達物質として分類されていますが、VIP 神経伝達物質であるボンベシン、ブラジキニン、バソプレシン、カルノシン、ニューロテンシン、ソマトスタチン、コレシストキニンについて不明であるのと同様、シナプス伝達におけるその役割は不明のままです。
- ノルアドレナリン - 最も重要な「覚醒メディエーター」の 1 つと考えられています。 ノルアドレナリン作動性投射は、上行網様体活性化システムに関与しています。 これは、脳幹の青みがかった斑点 (ラテン語の青斑核) と交感神経系の終末の両方の仲介者です。 CNS 内のノルアドレナリン作動性ニューロンの数は少ない (数千) が、脳内に非常に広い神経支配領域を持っています。
- ドーパミンは内部強化の化学因子の 1 つであり、喜び (または満足) の期待 (または期待) の感情を引き起こし、モチベーションのプロセスに影響を与えるため、脳の「報酬システム」の重要な部分として機能します。そして学ぶこと。
その他のモノアミン
- セロトニン - 中枢神経系で神経伝達物質の役割を果たします。 セロトニン作動性ニューロンは、脳幹、つまり橋核と縫線核にグループ化されています。 橋からは脊髄への下行投射があり、縫線核のニューロンは小脳、大脳辺縁系、大脳基底核、皮質へ上行投射を行っています。 同時に、背側縫線核と内側縫線核のニューロンは、形態学的、電気生理学的、神経支配の標的、および特定の神経毒物質(たとえばメタンフェタミン)に対する感受性において異なる軸索を生じます。
- ヒスタミン - ヒスタミンの一部は CNS に存在し、神経伝達物質 (または神経調節物質) の役割を果たすと考えられています。 一部の親油性ヒスタミン拮抗薬(血液脳関門を通過する抗ヒスタミン薬、たとえばジフェンヒドラミン)の鎮静作用は、中枢ヒスタミン受容体に対する遮断作用と関連している可能性があります。
その他の代表者
- アセチルコリン - 神経筋伝達を行うほか、副交感神経系の主要な神経伝達物質であり、神経伝達物質の中でコリンの唯一の誘導体です。
- アナンダミドは、痛み、うつ病、食欲、記憶、生殖機能のメカニズムに役割を果たす神経伝達物質および神経調節物質です。 また、虚血や再灌流による不整脈誘発効果に対する心臓の抵抗力も高めます。
- ATP (アデノシン三リン酸) - 神経伝達物質としての役割は明らかではありません。
- 血管作動性腸管ペプチド (VIP) - 神経伝達物質としての役割は明らかではありません。
- タウリン - シナプス伝達を阻害する神経伝達物質アミノ酸の役割を果たし、抗けいれん作用があり、また強心作用もあります。
- トリプタミン - トリプタミンは、哺乳類の脳において神経伝達物質および神経伝達物質としての役割を果たすと仮説が立てられています。
- エンドカンナビノイド - 細胞間シグナル伝達の役割において、それらはアセチルコリンやドーパミンなどの既知のモノアミン伝達物質と似ていますが、エンドカンナビノイドは多くの点でそれらとは異なります - たとえば、エンドカンナビノイドは逆行性シグナル伝達を使用します(シナプス後膜によって放出され、シナプス前膜に作用します)。 。 さらに、エンドカンナビノイドは親油性分子であり、水に溶けません。 それらは小胞に保管されていませんが、細胞の一部である膜二重層の不可欠な構成要素として存在します。 おそらく、それらは後で使用するために保存されるのではなく、「オンデマンド」で合成されます。
- N-アセチルアスパルチルグルタミン酸塩 (NAAG) は、哺乳類の神経系で 3 番目に豊富な神経伝達物質です。 これは、神経伝達物質の特徴的な特性をすべて備えています。ニューロンおよびシナプス小胞に集中し、活動電位の開始後にカルシウムの影響下で軸索末端から放出され、ペプチダーゼによる細胞外加水分解を受けます。 これは、グループ II 代謝指向性グルタミン酸受容体、特に mGluR3 受容体のアゴニストとして作用し、NAAG ペプチダーゼ (GCPII、GCPIII) によってシナプス間隙で親物質である NAA とグルタミン酸に切断されます。
- さらに、一部の脂肪酸誘導体 (エイコサノイドおよびアラキドン酸)、一部のプリンおよびピリミジン (アデニンなど)、および ATP については、神経伝達物質 (または神経調節) の役割が示されています。
アクション
神経伝達物質はホルモンと同様に主要なメッセンジャーですが、その放出と化学シナプスでの作用機構はホルモンとは大きく異なります。 シナプス前細胞では、神経伝達物質を含む小胞が神経伝達物質を局所的に非常に小さな体積のシナプス間隙に放出します。 放出された神経伝達物質は裂孔を通って拡散し、シナプス後膜の受容体に結合します。 拡散は遅いプロセスですが、シナプス前膜とシナプス後膜を隔てるような短い距離 (0.1 μm 以下) を通過するのに十分な速さで、ニューロン間またはニューロンと筋肉の間で迅速な信号伝達を可能にします。
神経伝達物質のいずれかが欠乏すると、さまざまな種類のうつ病などのさまざまな障害が引き起こされる可能性があります。
タバコやアルコールなどの薬物への依存の形成は、これらの物質の使用により、神経伝達物質セロトニンや、同様の神経伝達物質をブロック(置き換え)する他の神経伝達物質の生成メカニズムが活性化されるためであるとも考えられています。自然のメカニズム。
行動とメディエーター(アミノ酸)の関係のメカニズムのいくつかの説明は、「精神矯正の方法としてのニュートリツヴェティカ」という本に記載されています。
関連記事